
חזית המחקר
ממשק אוכלוסיות יתר של תנים – מתֵאוריה למציאות
דרור קפוטא המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקר
עידן טלמון המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקר
אלון רייכמן מחוז צפון, רשות הטבע והגנים
יהודה יהודה תחום בעלי חיים, מו"פ צפון מיג"ל, קריית שמונה
חווה גולדשטיין מחוז צפון, רשות הטבע והגנים
רוני קינג חטיבת מדע, רשות הטבע והגנים
דויד זלץ המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב

שועל מצוי הניזון מאשפה. גמלא, 2013 | צילום: עידו שקד
עמית דולב
מחוז צפון, רשות הטבע והגניםדרור קפוטא
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקרעידן טלמון
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקראלון רייכמן
מחוז צפון, רשות הטבע והגניםיהודה יהודה
תחום בעלי חיים, מו"פ צפון מיג"ל, קריית שמונהחווה גולדשטיין
מחוז צפון, רשות הטבע והגניםרוני קינג
חטיבת מדע, רשות הטבע והגניםדויד זלץ
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגבעמית דולב
מחוז צפון, רשות הטבע והגניםדרור קפוטא
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקרעידן טלמון
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגב, שדה בוקראלון רייכמן
מחוז צפון, רשות הטבע והגניםיהודה יהודה
תחום בעלי חיים, מו"פ צפון מיג"ל, קריית שמונהחווה גולדשטיין
מחוז צפון, רשות הטבע והגניםרוני קינג
חטיבת מדע, רשות הטבע והגניםדויד זלץ
המחלקה לאקולוגיה מדברית ע"ש מיטרני, אוניברסיטת בן-גוריון בנגבאוכלוסיות יתר של תנים מאיימות על יציבות המערכת הטבעית, גורמות נזקים לחקלאות, ומקֵלות על הפצת מחלת הכלבת. בשנות ה-90 חל גידול ניכר בטריפות של בקר וצאן בגולן, לצד ירידה חדה בגודל אוכלוסיות הצבאים. דילול נרחב של תנים בגולן לא הוביל לירידה בהיקפי הטריפה ולהתאוששות הצבאים. במחקר שלפניכם נבחנה השפעת ממשק מקורות המזון על אוכלוסיות התנים.
עבודת השדה כללה ספירות רכובות ומִשדור של 68 תנים, לצד דילול תנים וסילוק פגרי בקר מהמרעה. בשנים הראשונות, 2005–2010, בוצע דילול בלבד של 800–1,200 תנים לשנה, ולא ניכר שינוי בצפיפות התנים. לאחר הוספת ממשק הפחתת מזון (פעולות תברואה): א. חלה ירידה בהיקף דילול התנים בכ-40%. ב. לא ניכרה ירידה בצפיפות התנים. ג. גודל תחומי המחיה של תנים בקרבת יישובים חקלאיים היה קטן מאוד ביחס לשטחים פתוחים מרוחקים מיישוב. ד. מוקדי הפעילות הוסטו אל קִרבת היישובים החקלאיים. ה. ניכרה שונות בהיקפי טריפת עגלים בין משקים ובין שנים. ו. החלה מגמת גידול בספירות הצבאים בגולן.
מתוצאות המחקר עולה כי פעולות תברואה הן כלי חיוני לממשק אוכלוסיות התנים לצד פעילות משלימה של דילול, אולם היעדר טיפול משלים בצמצום מקורות מזון נוספים עלול לגרום ליעילות נמוכה בהקטנת אוכלוסיות היתר.
על קצה המזלג
- פעילות האדם ברמת הגולן הביאה להגדלת אוכלוסיית הטורפים (תנים וזאבים), ואלה גורמים לצמצום אוכלוסיית הצבאים ולנזקים לעדרי הבקר בגולן.
- מאחר שכבר הוכח כי שימוש בירי בלבד כשיטה לדילול אוכלוסיית התנים אינו מספק, יש למצוא ממשק יעיל יותר כדי להפחית את נזקי הטריפה.
- המחקר בחן את ההשפעה שיש לדילול באמצעות ירי בשילוב הפחתת מקורות המזון הזמינים, על גודל אוכלוסיית התנים ברמת הגולן, על הדינמיקה המרחבית שלהם ועל השפעת התנים על אוכלוסיות הנטרפים.
- הפחתת מקורות המזון הזמינים נמצאה, לפחות בטווח הזמן הקצר, כלא מספקת בצמצום אוכלוסיית התנים. יש לבחון אם יש גורמים (למשל חברתיים ומרחביים) המעכבים את השפעת הטיפול.
- כיום מתבסס הממשק למניעת נזקי טריפות של חיות בר, בין השאר, על תוצאות מחקר זה.
המערכת
הלכה למעשה
מאמר זה מתבסס על דו"ח מחקר מסכם שהוגש למשרד החקלאות ופיתוח הכפר ולרשות הטבע והגנים
מבוא
עד אמצע שנות ה-80 של המאה ה-20 נחשבו נזקי טריפת עגלים בצפון ישראל מועטים, אולם במהלך שנות ה-90 חל גידול ניכר בהיקפי הטריפה, ומרבית הנזקים התרכזו במרחב רמת הגולן. ב-1993 נאמדו נזקי הטריפה בגולן (מעל 20 משקים) ב-42,000 דולר [26]. בשנים 2010-1998 עמד היקף הטריפות בגולן על כ-300-100 פרטים של בקר וצאן (מעל 80% מהם בקר) [6]. טריפות אלה יוחסו בעיקר לזאב האפור (Canis lupus) ולתן הזהוב (Canis aureus) ביחסים של שני שלישים ושליש בהתאמה [6]. נזקי הטריפה התמקדו לרוב במרחב מצומצם, וגרמו לטריפה של עשרות עגלים בזמן קצר [6, 21, 26]. היקפי הטריפה הובילו לא אחת לפעולות דחק של חקלאים, שניסו להרעיל את הטורפים.
ממחקר שנועד לאמוד את גודל אוכלוסיית הזאבים בגולן ולהמליץ על דרכים להפחתת נזקי הטריפה [21] עולות הנקודות הבאות:
- גודל אוכלוסיית הזאבים בגולן עומד על כ-100-80 פרטים בשטח של פחות מ-1,000 קמ"ר. צפיפות זו גבוהה יחסית למוכר בעולם [21];
- תחום מחייתם קטן ממחצית מהמוכר באירופה (בישראל כ-46 קמ"ר [21], בדרום אירופה: 240-80 קמ"ר, בצפון אירופה: 414–500 קמ"ר) [17].
הדבר מלמד על היצע מזון רב, שמקטין את הצורך בתנועות ארוכות טווח. מחקר שבחן את הירידה בגודל אוכלוסיות הצבאים בגולן [5], הראה שאחד הגורמים המשמעותיים להיעדר התאוששות של הצבאים בגולן הוא קיומן של אוכלוסיות יתר של תנים. אוכלוסיות אלה מכונות מינים מתפרצים, כלומר אוכלוסיות שגדולות מכושר הנשיאה הטבעי של השטח, ושמתבססות על מקורות מזון זמין שמגיע מפעילות אנושית.
תובנות אלה הובילו להחלטה של רשות הטבע והגנים בראשית שנות ה-2000 לתת היתרי דילול לזאבים בהתאם לתכנית ממשק, ולקדם דילול נרחב של אוכלוסיות התנים. בד בבד, הוכנסו לשימוש כלבי הגנה וגדרות חשמליות, והוקמו חלקות המלטה ממוגנות לבקר [6]. למרות זאת, לא ניכרה הפחתה משמעותית בהיקף הטריפות עד לראשית הממשק המשולב בסוף שנת 2010, דבר המעיד על כך שאין באמצעים אלה מענה מספק.
בהתבסס על מחקרים קודמים [2, 21] עלתה ההשערה כי השארת פגרי בקר בשטח היא אחד הגורמים המשמעותיים לעודף מזון זמין לטורפים. במרחב הגולן עומד היקף גידול הבקר במרעה על כ-26,000 ראש, ולכן סביר שתמותה טבעית בשיעור של כ-5% תספק לאוכלוסיות הטורפים משאב מזון ניכר. הדבר מעלה את כושר הנשיאה של השטח, ומאפשר גידול יתר של אוכלוסיות הטורפים. מצב זה תורם להיווצרות אוכלוסיות יתר בצפיפות גבוהה של פי עשרה ויותר [1, 8, 12, 15]. נוסף על כך, היות שתנים הם אוכלי כול, אוכלוסיותיהם יכולות להיזון ממגוון מקורות מזון, כמו מזון לחיות משק, פֵּרות במטעים ושאריות פסולת [1, 8, 12, 20, 22]. אוכלוסיות יתר צפופות אלה תורמות לגידול בנזקי הטריפה לעדרי הבקר בהיקף של עד כ-250 פרטים בשנה ברמת הגולן [3, 6], לפגיעה באוכלוסיות הצבאים [5], לשיעור הפצה גבוה של כלבת [1] ולפגיעה בצינורות השקיה [1].
מחקר זה נועד לבחון את ההשפעה של הפחתה בזמינות מקורות מזון על אוכלוסיות התנים ועל אוכלוסיות הנטרפים – בקר וצבאים, בהתבסס על ההשערה, שהפחתה בזמינות מזון לטורפים צפויה להקטין את כושר הנשיאה, לצמצם את אוכלוסיותיהם בטווח הארוך [2, 8, 14] ולתרום להפחתת היקפי הטריפה.
שיטות
המחקר התבצע במרכז רמת הגולן ובדרומה בשטח של כ-100,000 דונם, הכולל חמישה יישובים חקלאיים (יונתן, רמת מגשימים, נטור, נוב ואבני איתן – איור 2ב) ושטחי מרעה עם כ-2,000 ראשי בקר. באזור המחקר קיימים מספר מקורות מזון אנתרופוגניים (התלויים בפעילות אדם, פירוט בנספח 1) המושכים אוכלוסיות תנים:
- פגרי בקר במרעה שיכולים להימצא בכל שטח המחקר;
- חקלאות בעלי חיים, הכוללת לולים, רפתות, דירים, מפטמות ומרכזי מזון. המזון הזמין בהם כולל כופתיות מזון לבעלי חיים, פגרים, שליות של פרות, תחמיץ וערֵמות זבל;
- פֵּרות במטעים ובכרמים (בעיקר בקיץ).
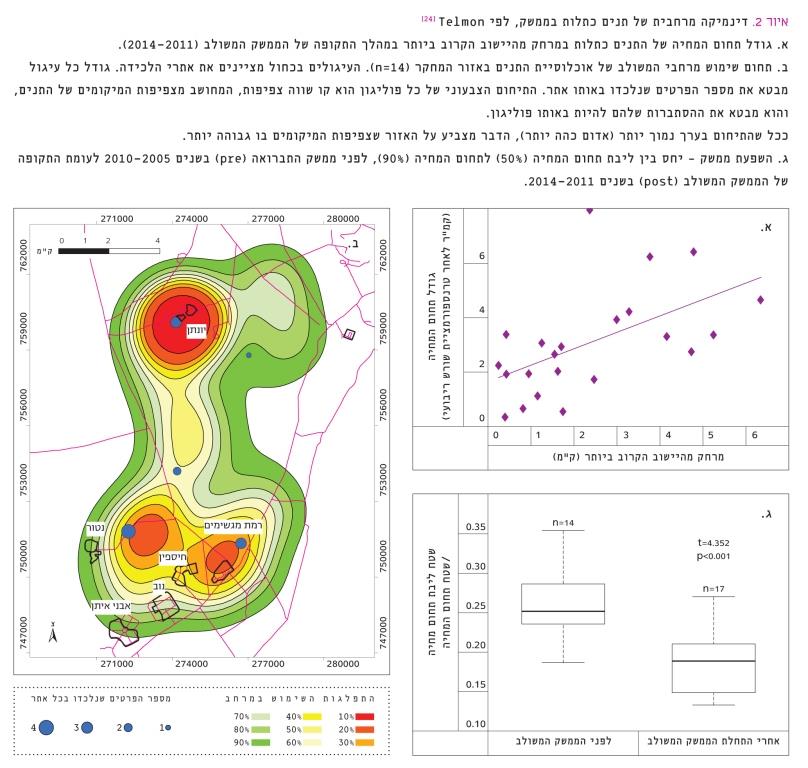
איור 2
דינמיקה מרחבית של תנים כתלות בממשק, לפי Telmon [24]
א. גודל תחום המחיה של התנים כתלות במרחק מהיישוב הקרוב ביותר במהלך התקופה של הממשק המשולב (2011–2014).
ב. תחום שימוש מרחבי המשולב של אוכלוסיית התנים באזור המחקר (n=14). העיגולים בכחול מציינים את אתרי הלכידה. גודל כל עיגול מבטא את מספר הפרטים שנלכדו באותו אתר. התיחום הצבעוני של כל פוליגון הוא קו שווה צפיפות, המחושב מצפיפות המיקומים של התנים, והוא מבטא את ההסתברות שלהם להיות באותו פוליגון.
ככל שהתיחום בערך נמוך יותר (אדום כהה יותר), הדבר מצביע על האזור שצפיפות המיקומים בו גבוהה יותר. ג. השפעת ממשק – יחס בין ליבת תחום המחיה (50%) לתחום המחיה (90%), לפני ממשק התברואה (pre) בשנים 2005–2010 לעומת התקופה של הממשק המשולב (post) בשנים 2011–2014 .
טיפולי ממשק
במחקר זה נבחנו שתי שיטות ממשק אפשריות לוויסות אוכלוסיות תנים: דילול על-ידי ירי מול הפחתת מזון שכללה סילוק פגרי בקר (פעולות תברואה). בשלב המקדים בוצעה ההשוואה בעזרת מודל מתמטי. מודל דמוגרפי זה של אוכלוסיות תנים התבסס על תוצאות קצבי רבייה, גיוס ושרידות, ללא פעולות ממשק שנאמדו במחקר קודם בגליל [2, 14] ובראשית מחקר זה (2010-2005). השיטות לאומדן מפורטות בהמשך. גיוס לאוכלוסייה אצל תנים מוגדר כתקופה שהגורים עוזבים את המסגרת המשפחתית [16], דבר שמתרחש לרוב בראשית החורף הראשון לאחר היוולדם.
בעזרת המודל הדמוגרפי בוצעה השוואה בין שתי שיטות הממשק המתארות את הדינמיקה של אוכלוסייה תחת היקפי דילול שונים או תחת הפחתת מזון ברמות שונות, כדי לאמוד את האפקטיביות הצפויה של שתי שיטות הממשק. הפרמטרים במודל של שיטות הממשק התבססו על נתוני אמת אמפיריים:
- מאוכלוסיית תנים בגליל, שם התבצע ניסוי סילוק מזון בלבד [2, 14];
- מאוכלוסיות תנים בגולן (מחקר זה בשנים 2005–2010), שם בוצע דילול תנים בלבד.
במחקר זה בגולן לא ניתן היה לבודד את ממשק הפחתת המזון כאמצעי לממשק התנים, ועל כן, ההשוואה של תוצאות אמפיריות למודל בוצעה בעזרת שני טיפולי ממשק כבסיס לבחינת השפעתם על הפחתת נזקי טריפה:
- דילול תנים בדרום הגולן – בוצע בשנים 2014–2015 על-ידי פקחי רשות הטבע והגנים, בוקרים וציידים, וכלל מאמץ ירי בתנים במהלך כל עונות השנה בניסיון לצמצם את אוכלוסיותיהם. שיעור הדילול השנתי (culling rate) חושב כיחס בין כמות התנים שדוללו לגודל האוכלוסייה המחושב בדרום הגולן.
- סילוק פגרי בקר (פעולות תברואה) משטחי מרעה – מראשית 2011 בוצע בכל מרחב רמת הגולן בעזרת פקח רשות הטבע והגנים וקבלן כילוי מטעם קרן נזקי טבע של משרד החקלאות (חברת ביואקולוגיה). מרבית הפגרים הועברו למתקן כילוי, וחלק מהם הועברו לתחנות האכלה לנשרים. נוסף על כך, בחוות גידול עופות באזור המחקר הוחלפו פחי הפגרים בפחים טמונים, שאינם נגישים לחיות בר.


מעקב אחר אוכלוסיות התנים
- הערכת הצפיפות התבססה על חמישה חתכים רכובים מוארי זרקור בנתיבים קבועים באורך ממוצע של כ-10 ק"מ כל אחד. החתכים התבצעו במרכז רמת הגולן ובדרומה, בסתיו ובאביב (בכל עונה 3–6 חזרות) בתחילת הלילה. הניתוח בוצע בשיטת Distance sampling [9].
- לימוד התנועה המרחבית של התנים התבסס על מִשדור עם קולרי VHF וקולרי GPS. חישוב תחומי מחיה בוצע בשיטת Kernel Density Estimation [20] באמצעות חבילת AdehabitatHR [10] בשפת R לאומדן תחומי מחיה [19].
- מדד אחידות השימוש בתחום מחיה משמש לתיאור היקף השימוש בתחום המחיה. הוא מבוסס על היחס בין ליבת תחום המחיה (isopleth 50%) לכל תחום המחיה (isopleth 90%) [22].
- הערכת שרידות התנים התבססה על חיישן תמותה בקולרים. ניתוח השרידות בוצע בעזרת תוכנת MARK [11] ושימוש במודולת known fates. בחינת המודלים המסבירים את המידע בצורה הטובה ביותר בוצעה בעזרת קריטריוני AIC המדרגים את המודלים שנבדקו.
מעקב אחר אוכלוסיות הנטרפים
- חישוב היקפי הטריפה של העגלים התבסס על בסיסי מידע ממדגם של משקים מייצגים ברמת הגולן.
- ניתוח הדינמיקה של גודל אוכלוסיות הצבאים במרחב המחקר התבסס על ספירת צבאים שנתית שמבצעת רשות הטבע והגנים [4].
מבנה המחקר
ממשקי הטיפול כללו את כל שטח המחקר, ונבחנה ההשפעה בין התקופה שכללה ממשק דילול בלבד (2005–2010), לבין התקופה שבוצעו בה ממשק משולב של דילול ופעולות תברואה (2011–2014).

תוצאות
מודל השוואה של גודל אוכלוסיות התנים כתלות בטיפולי ממשק
במודל המקדים (איור 1) נבחנו שתי שיטות ממשק לוויסות אוכלוסיות תנים: א. דילול על ידי ירי; ב. הפחתת מזון. המודל, המבוסס על נתוני אמת דמוגרפיים [2, 14], שימש בסיס לאמידת האפקטיביות של שתי שיטות הממשק. הנחת הבסיס הייתה שבעוד שדילול בירי משפיע ישירות על גודל האוכלוסייה, סילוק המזון משפיע עליה בעקיפין בלבד. ממשק דילול מקטין את האוכלוסייה, אך מגדיל את שיעורי השרידות והגיוס עקב הפחתת התחרות על משאבי המזון. אצל תנים מוגדרת עונת הגיוס לאוכלוסייה כתקופה שהגורים עוזבים את המסגרת המשפחתית [16]. הדבר מתרחש לרוב בעונת הרבייה העוקבת – בראשית החורף הראשון לאחר היוולדם. בכך מפצה האוכלוסייה על הדילול באופן מלא או חלקי. השפעת הדילול איננה לינֵארית, וייתכן שכשתבוצע במינון נמוך, קצב הגיוס יפצה על הדילול, והממשק לא יהיה אפקטיבי. לעומת זאת, ממשק הפחתת המזון גורם לעלייה בתחרות, ולכן צפוי להוביל לירידה בגודל האוכלוסייה עקב ירידה זמנית בקצבי הגידול, עד להתייצבותה על מספר פרטים נמוך יותר, התואם את כמות המזון החדשה. לפי תחזית המודל (איור 1), בממשק של דילול בירי בלבד, תקטן האוכלוסייה רק במעט גם כאשר פרופורציית הדילול תוגדל משמעותית. הדבר צפוי להיגרם בגלל שיעור הריבוי הגבוה יחסית של תנים (4–8 גורים בשגר [16]) שיפצו על הקטנת האוכלוסייה על-ידי גיוס ניכר בעונת הרבייה העוקבת. לפי המודל, רק לאחר שפרופורציית הדילול תעבור ערך סף של כ-70%, צפויה להיות ירידה משמעותית בגודל האוכלוסייה. ירידה חדה זו עלולה אף להוביל להכחדתה עקב קושי לשלוט בקצב הירידה. לפי המודל, בממשק של הקטנת מזון זמין בלבד (איור 1) צפויה אוכלוסיית התנים לקטון באופן לינֵארי ופרופורציוני לכמות המזון המסולקת, ללא תלות במאפיינים דמוגרפיים.
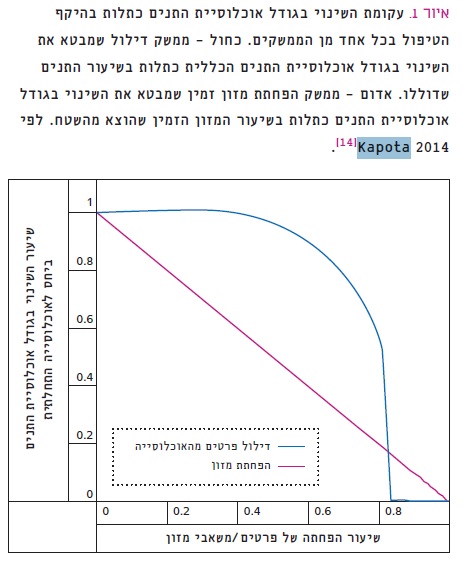
איור 1
עקומת השינוי בגודל אוכלוסיית התנים כתלות בהיקף הטיפול בכל אחד מן הממשקים.
כחול – ממשק דילול שמבטא את השינוי בגודל אוכלוסיית התנים הכללית כתלות בשיעור התנים שדוללו. אדום – ממשק הפחתת מזון זמין שמבטא את השינוי בגודל אוכלוסיית התנים כתלות בשיעור המזון הזמין שהוצא מהשטח.
לפי Kapota 2014 [14].
טיפולי ממשק
דילול תנים
ממשק דילול התנים בדרום הגולן התבצע בשטחים הפתוחים. הנחת הבסיס היא שמאמץ הדילול של כל המעורבים (פקחים, חקלאים וציידים) היה דומה במהלך השנים. בשנים 2012-2007 עמד היקף הדילול על 1,049.5±53.5 תנים בשנה (1,213, 1,178, 1,040, 808, 1,045 ו-1,013 בהתאמה). למרות הדילול הנרחב לא נראתה ירידה בהיקף הדילול בשנה עוקבת. שיעור הדילול המחושב בשנים 2006–2008 היה 0.35, 0.66 ו-0.59 בהתאמה. בשנים 2013–2014, שהיו שנים שלישית ורביעית בהתאמה להתחלת הממשק המשולב, נראתה ירידה של 47%-36 בהיקפי הדילול (674 ו-554 בהתאמה), מבלי שהיה שינוי במדיניות ההיתרים לדילול. הדבר עשוי ללמד על זמינות נמוכה יותר של תנים לדילול בשטחים הפתוחים לאחר מספר שנים של ממשק משולב.
סילוק פגרים (פעולות תברואה) משטחי מרעה
ממשק זה התבצע בהיקף מלא מאמצע שנת 2011. היקף הפגרים שפונו ברחבי רמת הגולן כולה בשנים 2011–2014 עמד על 975, 1,304, 1,130 ו-1,428 בהתאמה. אומדן של משקל הפגרים הכולל שפונו התבסס על ערך של 500 ק"ג לפרה ו-50 ק"ג לעגל. האומדן מלמד על היקף של 303±27.8 טונות פגרים לשנה בממוצע, וניכרת מגמת גידול עם השנים. נראה שפעילות זו תרמה לסילוק של מעל ל-80% מהפגרים הזמינים בעדרים המוסדרים.
בחינת ההשפעה של טיפולי הממשק על אוכלוסיות התנים
בוצע מעקב אחר 68 תנים שנלכדו ושהוצמד להם משדר. 14 תחת ממשק דילול בלבד (2007–2008), ו-54 תחת ממשק משולב (2010–2014). בממשק המשולב, ל-15 תנים הוצמד קולר עם ג'י-פי-אס.
צפיפות התנים
ניתוח רב-שנתי הראה שצפיפות התנים בדרום הגולן נאמדת ב-1–5 תנים לקמ"ר (נספח 2), ללא מגמת ירידה מתחילת ממשק הדילול (שנת 2005) והממשק המשולב (שנת 2011), וזאת למרות היקפי הדילול הנרחבים. ניכרה שונות עונתית רבה בין הסתיו לאביב (פי שניים לערך), שנובעת מההבדל במספר התנים לפני הגיוס ואחריו. ההבדל משתקף בשיעור הגיוס (נספח 2) שמחושב כאוכלוסיית הסתיו חלקי אוכלוסיית האביב שקדם לו.
שרידות וגיוס
ניתוח שרידות התנים הראה שהיא הייתה גבוהה וללא מגמת שינוי לאורך השנים (נספח 2) למרות היקפי הדילול הנרחבים. העדות היחידה לירידה בשרידות התנים תועדה בתחילת הממשק המשולב (דצמבר 2010–פברואר 2011) בחלק המזרחי של אזור המחקר (מרעה מבוא חמה) שמרוחק מיישובים. בחלק המערבי של אזור המחקר (מרעה רמת מגשימים), הסמוך ליישוב שלא בוצע בו ממשק סילוק מזון במתקני גידול, נותרה שרידות התנים גבוהה כמקודם. בחינת הקשר בין שרידות התנים (נספח 2) לשיעור הדילול הראתה שהשרידות נשארה גבוהה וללא שינוי גם כאשר שיעור הדילול עלה כמעט פי שניים, מ-0.35 ל-0.66.
למרות היעדר השינוי בשרידות, בחינת הקשר בין שיעור הגיוס (נספח 2) לשיעור הדילול מראה שניכרת תגובת פיצוי משמעותית על-ידי גיוס.
משמעות הדבר, שקצב הגיוס של אוכלוסיית התנים תלוי בצפיפות, בעוד ששרידות הבוגרים נותרה גבוהה וכמעט קבועה. הפיצוי על דילול האוכלוסייה מגיע, אפוא, מעלייה בגיוס הסתיו, ולא מעלייה בשרידות הבוגרים הנותרים. עובדה זו מצביעה על דרך ההשפעה של הירי על האוכלוסייה. פירוש הדבר שאין ירידה משמעותית בגודל אוכלוסיית הסתיו עם השנים, אבל ייתכן שמאמץ לדילול במהלך שנה, בין גיוס לגיוס, עשוי להשתלם בטווח הקצר ולהקטין את שיעור הטריפות.
דינמיקה מרחבית
גודל תחום המחיה (isopleth 90%) הממוצע של תנים היה 11±2.3 קמ"ר (n=22). נמצא קשר חיובי מובהק (רגרסיה לינֵארית, p=0.031) בין גודל תחום המחיה למרחק ליישוב הקרוב (איור 2א), שמשמעו שככל שמרכז תחום המחיה של התן קרוב יותר ליישוב, שטח תחום המחיה שלו קטן יותר (פחות מ-1 קמ"ר), ולהפך. תנים שמרכז תחום מחייתם היה קרוב ליישוב או בתוכו, לרוב לא התרחקו ממנו. דפוס השימוש המרחבי של אוכלוסיית תנים באזור המחקר (איור 2ב) מדגיש שהיישובים החקלאיים וחוות גידול בעלי החיים משמשים מוקד פעילות משמעותי של התנים. בחינת ההבדל בין גודל תחום המחיה של תנים לפני ואחרי תחילת הממשק המשולב לא הראתה הבדל מובהק (4W=110, p=0.73). ניתוח של מדד אחידות השימוש בתחום מחיה (היחס בין ליבת תחום המחיה (isopleth 50%) לכל תחום המחיה [isopleth 90%]) לפני ואחרי תחילת הממשק המשולב (איור 2ג), הראה כי לפני תחילת פעולות התברואה מדד האחידות היה גבוה בלמעלה מ-20% בהשוואה לערכו לאחר הממשק המשולב (מבחן t, t=4.3522, df=26.534, p<0.001). תוצאה זו מלמדת ששימוש התנים במרחב בתקופה של הממשק המשולב היה ממוקד יותר באזורים ספציפיים, לעומת התקופה של ממשק הדילול בלבד, שהשימוש בה היה מפוזר יותר במרחב.
בחינת ההשפעה של טיפולי הממשק על אוכלוסיות הנטרפים
היקפי טריפה של עגלים במרעה
מידע על היקפי טריפה של עגלים נאסף מ-16 משקי בקר שונים ברמת הגולן. לכל משק חושב היחס בין כמות העגלים שנטרפו לכמות העגלים שנולדו בכל שנה. היקף טריפות עגלים הראה מגמה מעורבת בין המשקים השונים, שונוּת בריכוזי פעילות הטורפים או הבדלים בהתמודדות המשקים השונים בתקופות שונות. בחינת ממוצע טריפות עגלים למשק לא הראתה מגמת שינוי מובהקת בתקופת המעקב (איור 3א). בשנים 2008–2011 (ממשק דילול) נראתה מגמת יציבות עם שיעור טריפה ממוצע של 3.4%. בשנים 2014-2012 (ממשק משולב) חלו ירידה בהיקפי הטריפה ב-2012–2013 (שיעור ממוצע של 2.1%), ולאחריה חזרה ב-2014 לשיעור טריפה הדומה לשנים קודמות.
דינמיקה של אוכלוסיות צבאים במרחב המחקר
ספירת הצבאים בדרום הגולן משמשת אמצעי נוסף לבחינת השפעת הטורפים על הנטרפים בשטחים הפתוחים. תוצאות המעקב שערכה רשות הטבע והגנים [4] מצביעות על יציבות בגודל האוכלוסיות בשנים 2003–2012 (איור 3ב). מ-2012 ואילך, כשנה וחצי לאחר תחילת הממשק המשולב, ניכרת מגמת גידול בגודל האוכלוסיות.
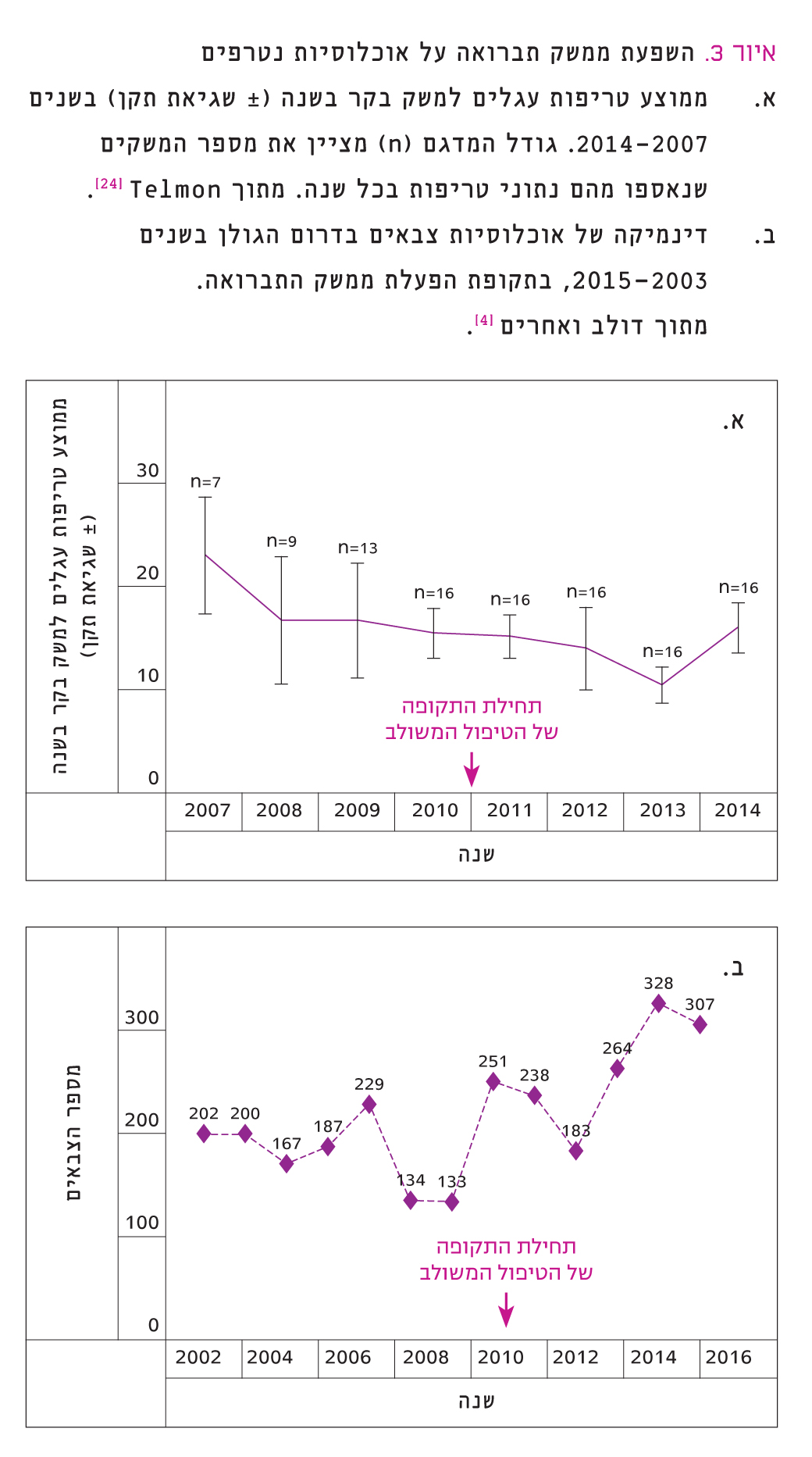
איור 3
השפעת ממשק תברואה על אוכלוסיות נטרפים
א. ממוצע טריפות עגלים למשק בקר בשנה (± שגיאת תקן) בשנים 2007–2014. גודל המדגם מציין את מספר המשקים שנאספו מהם נתוני טריפות בכל שנה. מתוך Telmon [24].
ב. דינמיקה של אוכלוסיות צבאים בדרום הגולן בשנים 2003–2015 , בתקופת הפעלת ממשק התברואה. מתוך דולב ואחרים [4].
דיון
בחינת ההשפעה של ממשק הדילול על אוכלוסיות התנים
בגלל יכולת הרבייה הגבוהה של תנים [16] נדרש דילול יעיל ובהיקף נרחב מהאוכלוסייה במרחב נתון כדי לגרום להפחתה משמעותית באוכלוסייה (איור 1). דילול בשיעור נמוך של האוכלוסייה יביא לרבייה מפצה, וכתוצאה מכך הדילול לא יהיה יעיל. הדמוגרפיה של התנים מאופיינת בשרידות קבועה וגבוהה של בוגרים, והגיוס תלוי צפיפות. היות שכך, השפעת הדילול תורגש באביב ותיעלם בעונת הסתיו. המאמץ הנדרש לדילול אינו לינֵארי מכמה סיבות: א. ככל שמאמץ הירי גדֵל, קשה יותר לפגוע בתנים עקב חשדנותם. ב. ככל שיש פחות תנים, קשה יותר לפגוע בהם. ג. הדילול מתבצע באזורים נגישים, ונותרים אזורים שהתנים יכולים להגר אליהם בתקופת דילול אינטנסיבי. יותר מכך, הדילול נותן פתרון זמני לאזור נתון, אולם גורם לעידוד שיעור הגיוס, ואיננו משפיע על השרידות של הפרטים הנותרים. מסיבה זו, בתקופה שהתבצע ממשק דילול בלבד, לא נצפה שינוי בצפיפות אוכלוסיות התנים, ולא היו ירידה בצפיפות התנים ובהיקפי הטריפה. בכל מקרה, נראה כי מאמץ הדילול הנחוץ להחזיק את אוכלוסיית הטורף בתחום הדרוש, גבוה עד כדי הפיכתו ללא יישומי.
בחינת ההשפעה של ממשק משולב (דילול ופעולות תברואה) על אוכלוסיות התנים
ניסוי מבוקר בגליל הראה שתגובתן של אוכלוסיות התנים לממשק תברואה בלבד מהירה, ועוצמתה גבוהה: השרידות פוחתת וההגירה עולה [2, 8, 14]. לפי המודל התאורטי (איור 1), כתוצאה מהממשק המשולב הייתה צפויה להתקבל ירידה משמעותית בצפיפות התנים ובשרידותם, ואולי גם ירידה בטריפות. בפועל אירעה ירידה בשרידות באחד האתרים (נספח 2) באזור המחקר שמרוחק מיישוב (כ-4 ק"מ דרומית מזרחית ליונתן, איור 2ב), שהצביעה על אפקטיביות ההשפעה של ממשק הפחתת המזון [3]. אף על פי כן, בקנה מידה נרחב של מרחב וזמן לא נראתה מגמה ברורה של שינוי. משמעות הדבר שיש עיכוב בהשפעת ממשק הפחתת המזון. מצב זה מפתיע, מאחר שמזון נחשב גורם מרכזי בוויסות אוכלוסיות [8, 12, 14, 18, 22, 24]. עם זאת, במצב של מזון עודף בכמות גדולה מאוד, עלולים להיות גורמים נוספים שאחראים על ויסות גודל האוכלוסייה, לדוגמה: חפיפה בתחומי מחיה, אינטראקציות חברתיות ועוד. מחקר שבחן עכברים מצויים (Mus musculus) הראה שבתנאי מרחב ומזון לא מוגבלים, האוכלוסייה מווסתת על-ידי אינטראקציות חברתיות [23]. מחקר אחר בגליל [1] הראה עדויות לכך שאוכלוסיית השועלים מסביב ליישובים חקלאיים שיש בהם לולים, מווסתת על-ידי גודל תחום המחיה ולא על-ידי מזון. משקלם של השועלים בתחומי היישובים, שרידותם ושיעורי הגיוס שלהם היו גבוהים יותר מאלה של השועלים בשטחים הפתוחים, בעוד ששיעור ההגירה שלהם היה נמוך [14]. משמעות הדבר, שאם אוכלוסיית התנים מווסתת על-ידי גורמים מרחביים או חברתיים, במצב של עודף מזון ניכר היא אכן צפויה להראות עיכוב בתגובה להפחתת המזון, כפי שנראה במחקר זה.
אי-פינוי של פגרי בקר משטחי מרעה לפני פעולות התברואה תרם לפיזור מזון רב בצורה אקראית בזמן ובמרחב. נראה כי פיזור זה גרם לכך שמדד השימוש בתחום המחיה היה גבוה אצל תנים ממושדרים לפני תחילת פעולות התברואה לעומת תנים ממושדרים אחרי תחילתן (איור 2ג), מאחר שלפני כן התנים השתמשו בתחום המחיה שלהם באופן נרחב יותר בגלל זמינות המזון במרחב. לאחר הפעלת ממשק התברואה, נראה שהתנים עברו להסתמך יותר על מקורות מזון קבועים ביישובים (איור 2ב), ועל כן המדד נמוך יותר. הסבר זה נתמך בתצפיות בתנים שמושדרו בקולר ג'י-פי-אס וסיפקו מידע מדויק בתדירות גבוהה על פעילות סביב מוקדי משיכה קבועים בשל מזון ממקור אנושי, כגון: יישובים, רפתות, לולים ומרכזי מזון לפרות.
לאור האמור לעיל, ובהסתמך על מחקרים קודמים [2, 8, 14, 24, 26] המצביעים על מזון כגורם משמעותי מאוד, נראה שבמקרה זה המצב מורכב עקב מגוון מקורות מזון נוספים, ועל כן ההשפעה של סילוק פגרי בקר בהיקף ניכר לא תרמה לשינוי בטווח זמן קצר. נראה כי צמצום הקונפליקט מצריך הרחבה של תהליך הפחתת מזון זמין ממקור אנושי למגוון רחב יותר של מוקדים (רפתות, דירים, מזבלות, מפטמות ומרכזי מזון) כדי להשיג את התוצאות הרצויות.
תודות
ברצוננו להודות לפקחי רשות הטבע והגנים במרחב ולבוקרי רמת הגולן על שיתוף הפעולה והעזרה בעבודת השדה ובאיסוף הנתונים, וליותם גנדלר על עזרתו בעבודת השדה.
- פעילות האדם ברמת הגולן הביאה להגדלת אוכלוסיית הטורפים (תנים וזאבים), ואלה גורמים לצמצום אוכלוסיית הצבאים ולנזקים לעדרי הבקר בגולן.
- מאחר שכבר הוכח כי שימוש בירי בלבד כשיטה לדילול אוכלוסיית התנים אינו מספק, יש למצוא ממשק יעיל יותר כדי להפחית את נזקי הטריפה.
- המחקר בחן את ההשפעה שיש לדילול באמצעות ירי בשילוב הפחתת מקורות המזון הזמינים, על גודל אוכלוסיית התנים ברמת הגולן, על הדינמיקה המרחבית שלהם ועל השפעת התנים על אוכלוסיות הנטרפים.
- הפחתת מקורות המזון הזמינים נמצאה, לפחות בטווח הזמן הקצר, כלא מספקת בצמצום אוכלוסיית התנים. יש לבחון אם יש גורמים (למשל חברתיים ומרחביים) המעכבים את השפעת הטיפול.
- כיום מתבסס הממשק למניעת נזקי טריפות של חיות בר, בין השאר, על תוצאות מחקר זה.
המערכת
נספחים
נספח 1. ריכוז מידע על מצב זמינות המזון לתנים בכל אחד מהיישובים שבאזור המחקר
להורדהנספח 2. דינמיקה רב-שנתית של אוכלוסיות התנים בדרום הגולן כתלות בממשק
להורדהמקורות
- דולב ע. 2006. מודל התפשטות מרחבי של מחלת הכלבת המבוסס על דינאמיקה של אוכלוסיות השועל המצוי בגליל, ככלי לפיתוח דגמי פיזור אופטימאליים של פתיונות חיסון כלבת. שדה בוקר: אוניברסיטת בן-גוריון בנגב.
- דולב ע, בינו ג, קארק ס ואחרים. 2013. השפעת סילוק מוסדר של פגרי עופות מלולים על אוכלוסיות טורפים ונטרפים בסביבה סמוכה. אקולוגיה וסביבה 4: 44–49.
- דולב ע, טלמון ע, קפוטא ד ואחרים. 2015. הפחתת נזקי טריפה של בקר במרעה – דו"ח מסכם. הוגש למדען משרד החקלאות ולרשות הטבע והגנים.
- דולב ע, סיני י, אורון ט ואחרים. 2015. סיכום ספירות צבאים במחוז צפון 2015. דו"ח פנימי, רשות הטבע והגנים.
- זלץ ד, קפלן ד, לוטן ר ודגני ר. 2002. בנית מודל טורף/נטרף לקביעת מדיניות ממשק לשם מניעת המשך התדרדרותה, שיקומה והבטחת יציבותה לאורך זמן של אוכלוסיית הצבאים ברמת הגולן. דו"ח מסכם למדען משרד המדע.
- רייכמן א. 2014. זאבים ומשק האדם בצפון ישראל דו"ח מסכם לשנת 2014. מסמך פנימי, רשות הטבע והגנים.
- Amador-Alcalá S, Naranjo EJ, and Jiménez-Ferrer G. 2013. Wildlife predation on livestock and poultry: Implications for predator conservation in the rainforest of south-east Mexico. Oryx 47: 243–250.
- Bino G, Dolev A, Yosha D, et al. 2010. Abrupt spatial and numerical responses of overabundant foxes to a reduction in anthropogenic resources. Journal of Applied Ecology 47:1262–1271.
- Buckland ST, Anderson D, Burnham K, et al. 2001. Introduction to distance sampling. Oxford University Press Oxford, UK.
- Calenge C. 2006. The package adehabitat for the R software: A tool for the analysis of space and habitat use by animals. Ecological Modelling 197: 516–519.
- Cooch E and White G. 2006. Program MARK: A gentle introduction. Colorado State University: Fort Collins.
- Fedriani JM, Fuller TK, and Sauvajot RM. 2001. Does availability of anthropogenic food enhance densities of omnivorous mammals? An example with coyotes in southern California. Ecography 24: 325–331.
- Holmern T, Nyahongo J, and Røskafta E. 2007. Livestock loss caused by predators outside the Serengeti National Park, Tanzania Tomas. Biological Conservation 135: 518–526.
- Kapota D. 2014. Demographic and movement responses of red foxes and Golden Jackals to spatial and temporal changes in food availability (Phd dissertatuin). Sede Boqer: Ben-Gurion University of the Negev.
- Lanszki J, Giannatos G, Dolev A, et al. 2010. Late autumn trophic flexibility of the golden jackal Canis aureus. Acta Theriologica 55: 361-370.
- Mendelson H and Yom-Tov Y. 1999. Fauna Palaestina: Mammalia of Israel. Jerusalem: The Israel Academy of Sciences and Humanities.
- Okarma H, Jedrzejewski W, Schmidt K, et al. 1998. Home ranges of wolves in Bialowieza Primeval forest, Poland, compared with other Eurasian populations. Journal of Mammalogy 79: 842-852.
- Oro D, Genovart M, Tavecchia G, et al. 2013. Ecological and evolutionary implications of food subsidies from humans. Ecology Letters 16: 1501–1514.
- R for Windows (Version 3.2.4). 2014. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
- Raichev EG, Tsunoda H, Newman C, et al. 2013. The reliance of the golden jackal (Canis aureus) on anthropogenic foods in winter in central Bulgaria. Mammal Study 38: 19–27.
- Reichmann A and Saltz D. 2005. The Golan wolves: The dynamics, behavioral ecology, and management of an endangered pest. Israel Journal of Zoology 51: 87–133.
- Rotem G, Berger H, King R, et al. 2011. The effect of anthropogenic resources on the space-use patterns of golden jackals. The Journal of Wildlife Management 75: 132–136.
- Strecker RL and Emlen JT. 1953. Regulatory mechanisms in house-mouse populations: The effect of limited food supply on a confined population. Ecology 34: 375-385.
- Telmon I. 2015. Movement ecology of an overabundant Golden jackal (Canis aureus) population in an environment rich with anthropogenic food resources (M.Sc thesis). Sede Boqer: Ben-Gurion University of the Negev.
- Worton B. 1989. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70: 164–168.
- Yom-Tov Y, Ashkenazi S, and Viner O. 1995. Cattle predation by the golden jackal Canis aureus in the Golan Heights, Israel. Biological Conservation 73: 19–22.



 מהדורה מודפסת
מהדורה מודפסת








