
חזית המחקר
שרידות יער אורנים על גבול המדבר בעקבות שנות בצורת קיצונית
אייל רוטנברג מכון ויצמן למדע, המחלקה למדעי כדור הארץ וכוכבי הלכת
ניר הר מחלקת יער, מרחב צפון, קק"ל ([email protected])
יצחק משה קרן קיימת לישראל, מרחב דרום
שמואל ספרינצין קרן קיימת לישראל, מרחב דרום
ז'וזה גרינצוויג הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושלים
דן יקיר המחלקה למדעי כדור הארץ וכוכבי הלכת, מכון ויצמן למדע ([email protected])

יקיר פרייזלר
המחלקה למדעי כדור הארץ וכוכבי הלכת, מכון ויצמן למדע; הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםאייל רוטנברג
מכון ויצמן למדע, המחלקה למדעי כדור הארץ וכוכבי הלכתניר הר
מחלקת יער, מרחב צפון, קק"ליצחק משה
קרן קיימת לישראל, מרחב דרוםשמואל ספרינצין
קרן קיימת לישראל, מרחב דרוםז'וזה גרינצוויג
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםדן יקיר
המחלקה למדעי כדור הארץ וכוכבי הלכת, מכון ויצמן למדעיקיר פרייזלר
המחלקה למדעי כדור הארץ וכוכבי הלכת, מכון ויצמן למדע; הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםאייל רוטנברג
מכון ויצמן למדע, המחלקה למדעי כדור הארץ וכוכבי הלכתניר הר
מחלקת יער, מרחב צפון, קק"ליצחק משה
קרן קיימת לישראל, מרחב דרוםשמואל ספרינצין
קרן קיימת לישראל, מרחב דרוםז'וזה גרינצוויג
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםדן יקיר
המחלקה למדעי כדור הארץ וכוכבי הלכת, מכון ויצמן למדעתמותת עצים נרחבת נצפתה בקיץ 2010 ביער יתיר, בגבול תפוצת עצי היער, לאחר עשור שחמש שנים מתוכו שנות בצורת, וששתיים מהן רצופות. התמותה התאפיינה בתבנית של כתמי עצים מתים לצד עצים חיים. תבנית התמותה הובילה למחקר השוואתי ביער בין החלקה והעץ לשם זיהוי הגורמים המסבירים תופעה זו. מתוצאות המחקר עולה, כי לתנאי בית הגידול – של העץ הבודד או של קבוצת עצים – ישנה חשיבות גדולה לשרידות העצים בתנאי יובש; מיקום העץ בקרקע רדודה, סלעית ואבנית על בסיס סלע קירטון רך עדיף על מיקום בקרקע עמוקה על בסיס אבן גיר. בית הגידול המיטבי מאפשר שימור של לחות הקרקע, וכתוצאה מכך, את צבירת הביומסה ההכרחית לשרידות העץ. בחינה ממושכת של עצים סמוכים ברמות עקה שונות הראתה שתנאי בית הגידול משקפים הבדלים הן בפוטנציאלי מים שנמדדו טרום שחר הן בקצב גדילת העצים, דבר המצביע על ההטרוגניות בתת-הקרקע, שמובילה לתגובה שונה של עצים סמוכים לאותם תנאי אקלים.
התוצאות מצביעות על כך שצפיפות העומד הממוצעת היא גורם משני מבחינת יכולות היער והתמודדותו עם בצורות, וכי השונות הגבוהה בתנאי בית הגידול מובילה ליכולת התמודדות שונה של העץ הבודד או קבוצת עצים עם אותם תנאי אקלים. בבואנו לבחון את משטר הטיפול ומדיניות הנטיעות העתידיות וכן את מצב היערות הנטועים באזורים שחונים, ניתן להשתמש בכלים שנרכשו במסגרת מחקר זה ולבחון את עמידותם של עומדי העצים תוך שימוש בהטרוגניות בתוך החלקה, ובעקבות הבנת ההטרוגניות – את התגובות השונות לבצורת של עצים שכנים.
על קצה המזלג
- תמותת עצים כתוצאה משינוי האקלים היא בעיה שיערנים מתמודדים איתה בכל העולם. בארץ, יערות אורנים על ספר המדבר רגישים במיוחד לשינוי האקלים.
- במחקר נבחנו הסיבות לתמותה נרחבת של עצי אורן ירושלים ביער יתיר לאחר מספר שנות בצורת, לנוכח תצפיות של פיזור לא אחיד במרחב של העצים המתים.
- מסתבר שמאפיינים מקומיים של בית הגידול – במיוחד המסלע והקרקע – משפיעים במידה רבה על זמינות המים לשורשי העצים, ומכאן על סיכויי ההתפתחות והשרידות של עצי אורן ירושלים באזורים יובשניים למחצה, ואילו לצפיפות החלקה אין השפעה משמעותית.
- במאמר מוצעת בחינה גאומורפולוגית פשוטה של היער, המאפשרת זיהוי של האזורים הרגישים לתמותה.
- תוצאות המחקר מקדמות את ההבנה של הגורמים הקשורים לתמותת עצים ביערות, ומציעות למנהלי יערות באזורים יובשניים למחצה כלי שצריך להנחות פעולות ממשק ביער, בייחוד דילולים ונטיעות.
המערכת
הלכה למעשה
מבוא
תצפיות ברחבי העולם מצביעות על עלייה בתכיפות ובחומרה של בצורות בעשורים האחרונים בד בבד עם עלייה בתמותת יערות [6, 12], המיוחסת בחלקה לעלייה בטמפרטורת האוויר, למיעוט משקעים ולשינוי במשטר הגשמים. שינויים אלה מפֵרים את מאפייני בית הגידול ויחסי הגומלין שבין העץ לסביבתו, וגורמים לשינויים בתכולת המים בקרקע, להגברת קצב ההתאדות, להתפתחות חרקים המזיקים לצמח ועוד [5 ,7, 8, 10]. מאמץ מחקרי אקו-פיזיולוגי באזורי אקלים שונים בעולם מבוצע כעת לצורך הבנת תגובת היערות לבצורות ממושכות ולהבנת הסיבות והגורמים לתמותה [7, 17]. כמו במקומות אחרים בעולם, נצפתה ברחבי ישראל תמותה מוגברת ביערות אורן נטועים ובוגרים במהלך שנת 2010 בעקבות מספר שנות בצורת [1]. יערות האורן מהווים כ-40% משטח היערות בישראל, ומשתרעים על כחצי מיליון דונם [3], ואורן ירושלים (Pinus halepensis) הוא המין העיקרי בהם.
נמצא שלאורן ירושלים יתרונות רבים בהתבססות בקרקעות הנחשבות קשות לנטיעה [11], ועמידות והתאמה לאזורים יובשניים [9, 13]. בזכות תכונות אלה היה המין הדומיננטי בנטיעות קק"ל עד שנות ה-80 [11]. מחקרים קודמים על תמותת אורן ירושלים ביערות דרומיים (יתיר, להבים, להב וכרמים) הראו כי ניתן לחזות את תמותת העצים על פי קצב הגידול של טבעות הגדילה השנתיות, אך לא נעשה קישור בין שרידות העצים לבית הגידול המקומי של העץ הבודד. במקרה זה, שרידות העצים הללו שויכה להבדלים גנטיים ופיזיולוגיים בין העצים (ביער כרמים) [4], אף על פי שלשונוּת המרחבית, להרכב הקרקע ולסוג הקרקע ישנה השפעה רבה על חיוניות העצים באזורים צחיחים, ולפיכך יש להם משקל משמעותי בבחירת מין עצים לנטיעה ובבחירת בית הגידול [11].
מחקר זה בא לבחון את הסיבות והגורמים שהובילו לתמותת עצים נרחבת ביער יתיר במהלך שנת 2010. יער יתיר ניטע לפני כ-50 שנה, והוא היער הנטוע הגדול בישראל. הוא משתרע על כ-30,000 דונמים, ממוצע המשקעים הרב-שנתי בו (±86)276 מ"מ, ואורן ירושלים הוא המין הדומיננטי בו.
בין השנים 2000-2010 פקדו את אזור יער יתיר חמש שנות בצורת (על פי מדד SPI Standardized Precipitation Index), שמתוכן שנתיים חמורות ורציפות (עם 340 ימים ללא גשם בין 2008–2009). לאחר שנות בצורת חמורות אלה נצפתה בשנת 2010 תמותת עצים שלא נצפתה ב-45 שנות קיום היער. מהערכה המבוססת על צילומי אוויר מקיץ 2010 זוהו כ-50,000 עצים מתים ברחבי היער. את תשומת לב החוקרים משכה העובדה שפיזור העצים המתים לא היה אחיד, ושהתמותה אופיינה בכתמים של קבוצת עצים מתים בצמידות לקבוצת עצים חיים. בעקבות זאת, התמקד המחקר בהשוואה בין כתמי התמותה לעצי היער סביבם.
במחקר בחנו מספר גורמים שהיו עשויים להשפיע על התמותה: השפעת התבנית הגאומורפולוגית על מצב העומד, ההבדלים הגנטיים בין העצים, הקשר שבין תכונות הקרקע ותת-הקרקע לתבניות התמותה ומחקר פיזיולוגי של העץ הבודד. הנחת המחקר הייתה כי לשילוב גורמים אלה ישנה השפעה על חיוניות העצים ועל שרידותם בשנות בצורת קיצוניות.
מספר השערות מחקר הובילו את תכנון המחקר וביצועו ונבחנו בו:
- ישנם הבדלים גנטיים בין העצים השורדים לעצים המתים
- כתמיות התמותה נובעת מההבדלים הגאומורפולוגיים בין תאי שטח השונים
- שיפוע, מפנה, גיל וצפיפות החלקה הם גורמים משמעותיים בסיכויי הישרדות העצים
- מבנה תת-הקרקע של החלקות השורדות שונה משל החלקות הרגישות
השערות אלה נבחנו לאורך שלבי המחקר השונים תוך שלילה ואישוש של השערות לאורך הדרך, כפי שיוצג בהמשך.
שיטות וחומרים
בעקבות התמותה הזמינה קק"ל בשנת 2010 צילומי אוויר של היער (אקרוס ובניו בע"מ), ולאחר מכן פותח אלגוריתם לזיהוי העצים המתים ולמיקומם, ששולב כשכבת ממ"ג (GIS) שכללה מיקום של כל אחד מהעצים המתים ביער (איור 1 בנספח). האלגוריתם אומת וכויל בשטח (מידת התאמה מעל 90%), ונראה שפיזור העצים המתים הוא כתמי: קבוצת עצים מתים ליד עצי היער החיים (איור 2 בנספח). מתוך כלל העצים המתים סומנו בשטח 20 חלקות (בכל חלקה אותרה תת-חלקה של עצים מתים – 'חלקה רגישה', ולידה תת-חלקה חיה – 'חלקה עמידה') המפוזרות באופן מייצג ברחבי היער, עם אותם תנאים מרחביים (שיפוע, מפנה, נוחות הגעה לרכבי שטח ועבודה). חלקה (שטח של 800 מ"ר) הוגדרה רגישה או עמידה כאשר יותר מ-80% מהעצים בשטחה נמצאו מתים או חיים, בהתאמה. המחקר התמקד בהשוואה בין חלקות משני סוגים אלה, בסך הכול 40 תתי-חלקות מעקב, הכוללות 20 תתי-חלקות מכל סוג.
בחינת קצב גדילת העצים בחלקות השונות
במהלך שנת 2011 נערך סקר מצאי סטנדרטי (צפיפות, קוטר, גובה, כיסוי צמרות, חיוניות עצים), על פי פרוטוקול ושיטות המדידה של קק"ל, בכל תתי-ההחלקות ('רגישות' ו'עמידות'), בתוספת ספירת גדמים לצורך הערכת הצפיפות הקודמת. נוסף על כך, צוינו גיל החלקה, השיפוע והמפנה. הביומסה העל-קרקעית של העצים בחלקות אלה חושבה בעזרת משוואות אלומטריות, שפותחו לתנאי יער יתיר על-ידי Grünzweig et al [14]. נוסף על כך, נמדד קצב גידול הטבעות השנתיות באמצעות קדוחות (בעצים חיים) ופרוסות עצים (בעצים מתים) ב-11 מתוך 20 חלקות המחקר (באמצעות תוכנת Windendro). במחקר אחר, הנערך על-ידי המחברים, מנוטר קצב הגידול של 30 עצים משנת 2001 בתחנת המחקר של מכון ויצמן, הממוקמת במרכז היער. המדידה מתבצעת בעזרת סרט מדידה המחובר לקפיץ (דנדרומטר), ותוצאות הניטור מוצגות להלן (איור 1).
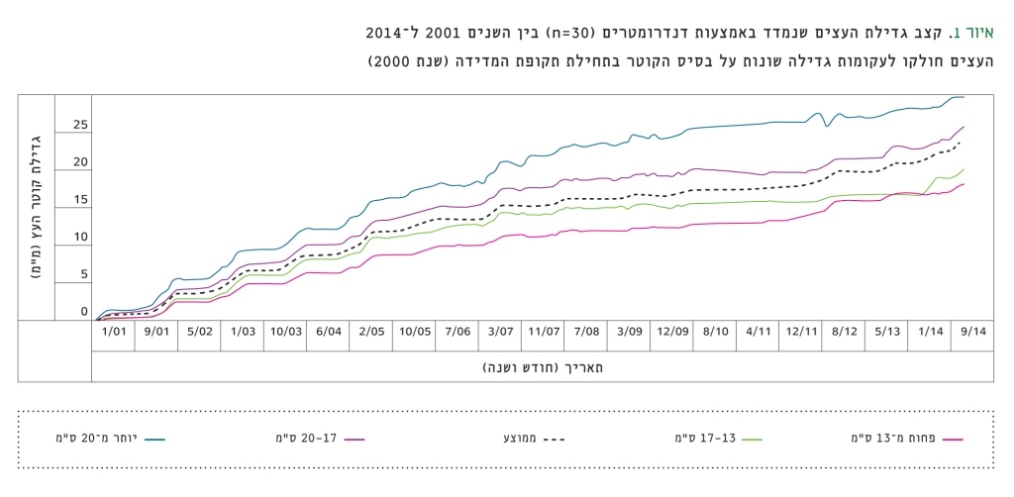
איור 1
קצב גדילת העצים שנמדד באמצעות דנדרומטרים (n=30) בין השנים 2001 ל-2014
העצים חולקו לעקומות גדילה שונות על בסיס הקוטר בתחילת תקופת המדידה (שנת 2000)
בחינת כיסוי הקרקע, אפיון תת-הקרקע והשורשים
ב-11 מתוך 20 חלקות המחקר נערך סקר קרקע מקיף באמצעות חפירת תעלה באורך של 7–10 מטרים ובעומק של 1–1.5 מטר. הסקר כלל בדיקות קרקע כימיות ופיזיקליות ובדיקות של אחוז האבנים, קוטר השורשים, פיזורם וצפיפותם לאורך פרופיל עומק הקרקע בשלושה מקטעי מדידה לאורך התעלה (איור 2). החפירה מוקמה במרכז כל חלקה, והניתוח הסטטיסטי נערך על-ידי השוואה מזווגת של כל צמד חלקות ('רגישות' מול 'עמידות').

איור 2
צוותי המחקר בתעלות שנערך בהן סקר הקרקע בחלקות המתות (למעלה) והחיות (במרכז)
סקר שורשים נערך לאורך מקטעי המדידה בעזרת מסגרת מדידה (למטה).
נוסף על כך, בדיקת כיסוי הקרקע (בחינת נוכחות סלע או קרקע ב-15 ס"מ העליונים) נערכה בחלקות אלה ב-36 נקודות דגימה לדונם, סך הכול 396 דגימות לכל סוג חלקה.
כדי לבחון את הקשר בין כיסוי הקרקע לבין התמותה בקנה המידה של כלל היער נותחו צילומי אוויר משנת 1945 (טרום היער), המאפשרים לזהות את כיסוי הקרקע ולאפיינו לאזורים סלעיים ולאזורים בעלי קרקע עמוקה, שזוהתה על פי קיום פעילות חקלאית. צילומים אלה הוצלבו עם שכבת התמותה (ArcMap 10.0) לצורך קישור בין כיסוי הקרקע לצפיפות התמותה ליחידת שטח.
מחקר פיזיולוגי של העץ הבודד
בחמש מתוך עשרים חלקות המחקר נערך גם מחקר פיזיולוגי לעצים, שבמסגרתו סווגו עצים על פי רמת העקה שהם שרויים בה. הסיווג הראשוני התבסס על מצבם החיצוני של העצים (phenotype) תוך שימוש בצבע המחטים ובאורכן כמדד העיקרי. על פי הסיווג הוגדר מדד חיוניות בסקאלה של 0–4: 4 הוא העץ החיוני ביותר, ו-0 מייצג עץ מת. נבחרו עצים שכנים בכל חלקה, המייצגים רמות עקה אלה (חמישה עצים בחלקה, בסך הכול 25 עצים). בהמשך נבחן הסיווג על-ידי מספר מדדים כמותיים, פיזיולוגיים ומורפולוגיים, שנמדדו בשטח ובמעבדה וכללו: פוטנציאל מים טרום שחר המייצג את פוטנציאל המים בקרקע (תא לחץ – Arimad), חילופי גזים, מדידת אורך המחטים השנתיות, אחוז החנקן והרכב האיזוטופים, ומדידת עובי הטבעות השנתיות. חלק מתוצאות המחקר מוצגות כאן (להרחבה ולפירוט פרק השיטות והשיטות הסטטיסטיות ראו נספח).
תוצאות
בבחינת ההבדלים בין חלקות רגישות וחלקות עמידות, נמצא כי הקוטר, הגובה והביומסה של העצים בחלקות ה'עמידות' היו גבוהים באופן מובהק מאלה שבחלקות ה'רגישות', וזאת בהשוואה לממוצע צפיפות העצים וגיל החלקה, שהיו זהים (טבלה 1). מתוצאות סקר המצאי בחלקות המחקר נמצא כי כ-73% מהעצים המתים היו בעלי קוטר קטן מ-17 ס"מ, וקוטרם של 52% מהעצים החיים היה מעל 17 ס"מ. ממוצע הביומסה העל-קרקעית של העצים החיים היה גבוה באופן מובהק משל העצים המתים, עם ±1.5123 לעומת ±160.5 ק"ג לעץ בהתאמה (P<0.001).

טבלה 1
ערכים ממוצעים ושגיאת התקן (בסוגריים) של תוצאות סקר המצאי
התוצאות מייצגות ממוצע של 20 חלקות חיות ו-20 יבשות. צפיפות הנטיעה (בשורה השנייה) מוערכת על פי מספר הגדמים שנמצאו במדגם. כוכביות (*) מסמנות הבדלים מובהקים בין החלקות. (*= רמת מובהקות p<0.05, **=רמת מובהקות p<0.005).
נוסף על כך, ממדידת קצב גדילת היקף העץ, הנמדד ביער ברציפות משנת 2001, עולה כי סך הגדילה המצטברת הממוצעת ב-13 השנים האחרונות (2001–2013) של עצים שקוטרם בתחילת תקופת המדידה היה קטן מ-17 ס"מ, היה נמוך בכ-50% מאשר עצים שקוטרם היה גדול מ-17 ס"מ (איור 1), כך שקוטר העצים נמצא כמצביע עיקרי על סיכויי שרידות עצים.
בחינת הקשר בין צפיפות החלקה לקוטר העצים הראתה קשר מתון בין הירידה בצפיפות לקוטר העצים. קשר זה נמצא חזק יותר בחלקות ה'עמידות' לעומת ה'רגישות' (R2=0.56 ו-0.36, כפי שמוצג באיור 3 בנספח).
השוואת עובי הטבעות השנתיות בין צמדי העצים החיים והיבשים מראה שונות משמעותית בדפוס הגדילה של העצים השכנים. החל בשנות ה-80 היה קצב הגדילה של העצים שמתו ב-2010, קטן ב-54±2% מזה של שכניהם החיים (איור 3א), והבדל זה החל להתבטא כבר משנת 1974. שנת הבצורת של 1984 הגדילה את הפער בין צמדי העצים, ובסופו של דבר הובילה לתמותה הסופית של העצים המוחלשים אחרי 25 שנים.
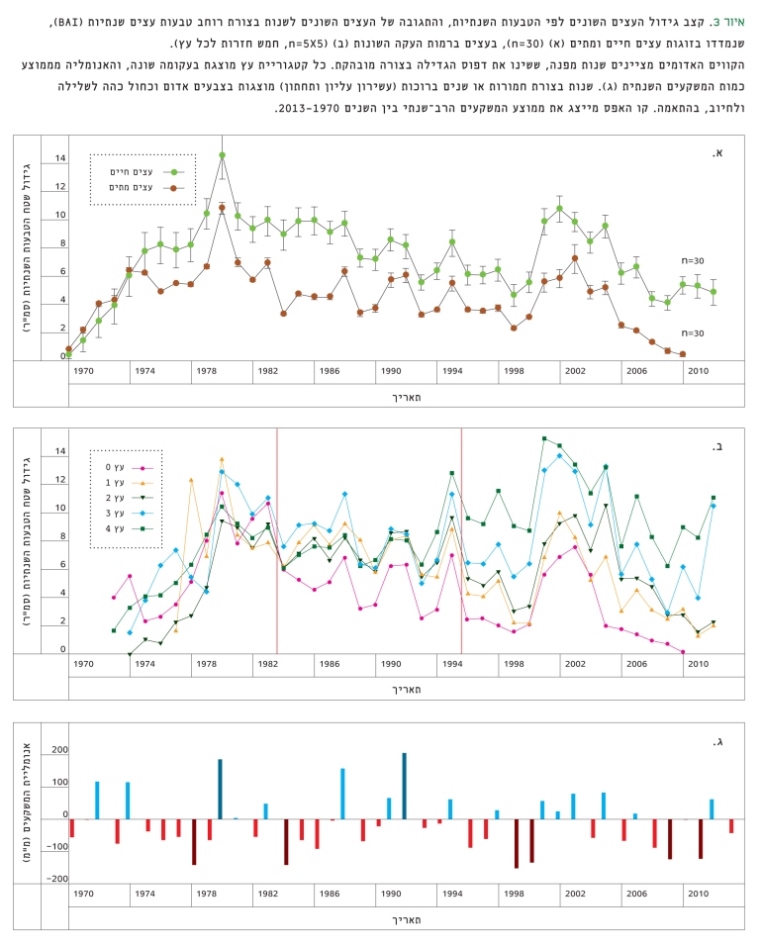
איור 3
קצב גידול העצים השונים לפי הטבעות השנתיות, והתגובה של העצים השונים לשנות בצורת רוחב טבעות עצים שנתיות (BAI), שנמדדו בזוגות עצים חיים ומתים (א) (n=30), בעצים ברמות העקה השונות (ב) (n=5X5, חמש חזרות לכל עץ).
הקווים האדומים מציינים שנות מפנה, ששינו את דפוס הגדילה בצורה מובהקת. כל קטגוריית עץ מוצגת בעקומה שונה, והאנומליה מממוצע כמות המשקעים השנתית (ג). שנות בצורת חמורות או שנים ברוכות (עשירון עליון ותחתון) מוצגות בצבעים אדום וכחול כהה לשלילה ולחיוב, בהתאמה. קו האפס מייצג את ממוצע המשקעים הרב-שנתי בין השנים 2013-1970 .
כיסוי הקרקע, אפיון תת-הקרקע והשורשים
החלקות ה'רגישות' התאפיינו בקרקע עמוקה יחסית (107±13 ס"מ), עם מעט סידן (5±0.7 מילי-אקוויוולנט לליטר) ועם תצבירי גיר בעומק, לעומת החלקות ה'עמידות' שהתאפיינו בקרקע רדודה יחסית (75±11 ס"מ), סלעית ואבנית (איור 4) עם סלע קירטון רך כסלע אם, ועם ריכוז סידן גבוה יותר (9.5±2 מילי-אקוויוולנט לליטר, p<0.05).
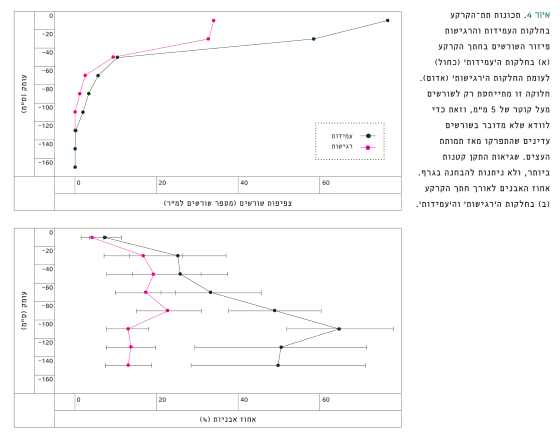
איור 4
תכונות תת-הקרקע בחלקות העמידות והרגישות פיזור השורשים בחתך הקרקע (א) בחלקות ה'עמידות' (כחול) לעומת החלקות ה'רגישות' (אדום). חלוקה זו מתייחסת רק לשורשים מעל קוטר של 5 מ"מ, וזאת כדי לוודא שלא מדובר בשורשים עדינים שהתפרקו מאז תמותת העצים. שגיאות התקן קטנות ביותר, ולא ניתנות להבחנה בגרף. אחוז האבנים לאורך חתך הקרקע (ב) בחלקות ה'רגישות' וה'עמידות'.
בחינת כיסוי פני הקרקע (Cs), הראתה כי בחלקות ה'עמידות' אחוז הסלע היה 36±7% ובחלקות ה'רגישות' רק 8±3%, p=0.004 (טבלה 1). צפיפות השורשים בפרופיל הקרקע, מפני הקרקע עד עומק של 1 מטר, הייתה גבוהה משמעותית, עד פי שניים, בחלקות ה'עמידות' לעומת החלקות ה'רגישות' (איור 4).
מבחינה מרחבית וגאומורפולוגית, על-ידי שימוש בצילומי אוויר ישנים וסיורי שטח, נראה כי בתי גידול בעלי הסתברות גבוהה יותר לתמותה, שהתאפיינו בין השאר בקרקע עמוקה, נמצאו בעמקים רחבים ובהם טרסות מעשה אדם שאוספות נגר, באוכפים, בקווי רכס ובשרידי טרסות מעשה אדם במדרונות (איור 5 ואיור 2 בנספח). בתי גידול סלעיים שנמצאו פחות רגישים לתמותה ונמצאו בעיקר בכיפות, בקווי רכס ובמדרונות סלעיים.

איור 5
דוגמאות לתופעת התמותה באזורים קרקעיים לעומת שרידות העצים בקרקע סלעית במקום שאין עצים בתמונה היו עצים מתים.
מאפייני הפיזיולוגיה של העץ הבודד
סיווג רמת העקה על פי מצבו החיצוני של העץ נמצא קשור באופן מובהק p<0.05)) לשלושה מדדים כמותיים: פוטנציאל המים טרום שחר, אורך המחטים ורוחב הטבעות השנתיות, שיפורטו להלן. פירוט נוסף של העצים וההבדלים החיצוניים מופיע בטבלה 2.
פוטנציאלי המים שנמדדו טרום שחר בין העצים ברמות העקה השונות, הראו הבדל מובהק (p<0.05) בכל העונות, למעט בשיא החורף. בקיץ 2012 ובאביב וקיץ 2013 נמצאו ההבדלים המשמעותיים ביותר בין העצים השונים (איור 6).
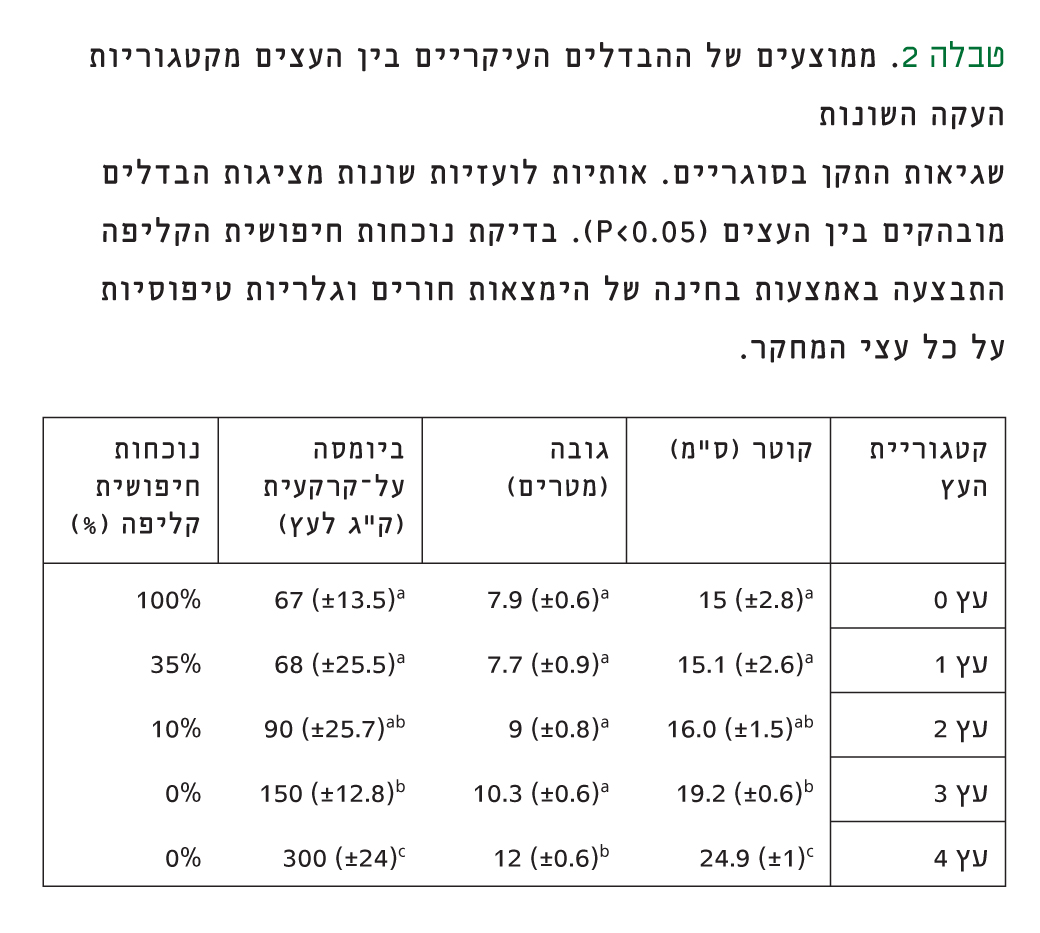
טבלה 2
ממוצעים של ההבדלים העיקריים בין העצים מקטגוריות העקה השונות
שגיאות התקן בסוגריים. אותיות לועזיות שונות מציגות הבדלים מובהקים בין העצים (P<0.05). בדיקת נוכחות חיפושית הקליפה התבצעה באמצעות בחינה של הימצאות חורים וגלריות טיפוסיות על כל עצי המחקר.
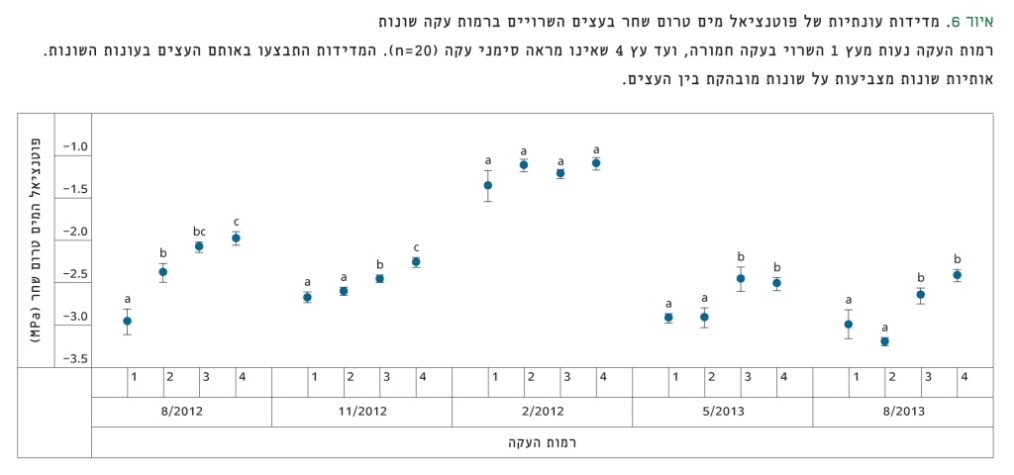
איור 6
מדידות עונתיות של פוטנציאל מים טרום שחר בעצים השרויים ברמות עקה שונות
רמות העקה נעות מעץ 1 השרוי בעקה חמורה, ועד עץ 4 שאינו מראה סימני עקה (n=20). המדידות התבצעו באותם העצים בעונות השונות. אותיות שונות מצביעות על שונות מובהקת בין העצים.
אורך המחטים שגדלו ב-3–4 השנים האחרונות, הראה היררכיה ברורה בין העצים ברמות העקה השונות. בכל השנים מחטי העצים החיוניים (3 ו-4) היו ארוכות ב-1.5 ס"מ בממוצע ממחטי עצים מקטגוריה '2' וב-2.5 ס"מ ממחטי העצים מקטגוריה '1' (p<0.05). נוסף על כך, תגובת העצים השונים לכמות המשקעים הייתה שונה, והדבר התבטא באורך המחטים. העצים המוחלשים (1 ו-2) הראו רגישות גבוהה לכמות המשקעים השנתית (r2=0.87, 0.92 בהתאמה) לעומת העצים החיוניים (3 ו-4) שהראו תגובה מופחתת (0.52 ו-0.65= r2) (איור 4 בנספח).
בהשוואת רוחב הטבעות השנתיות בין העצים ברמות העקה השונות, נמצא כי החל בשנת 1984 קטן קצב הגדילה הממוצע של העצים שהתייבשו ב-2010 (מסוג 0) ב-46±0.1% בהשוואה לעצים ששרדו (1-4). הפיצול הראשון בקצב הגדילה בין העצים חל לאחר שנת בצורת חמורה (146 מ"מ, כמחצית מהממוצע הרב-שנתי), שלאחריה שנתיים רצופות עם כמות משקעים נמוכה. משנת 1996 אובחנה התבדלות נוספת בקצב הגדילה בין העצים השורדים, בהתאמה לרמות העקה הנצפות כיום. קצב הגדילה של העצים השרויים בעקה (1 ו-2) היה קטן ב-29±0.05% מקצב הגדילה של העצים ללא עקה (3 ו-4). פיצול זה נוצר לאחר שנות הבצורת 1995–1996 (איור 3ב, איור 3ג). שינוי נוסף בקצב גדילת הטבעות התרחש בעקבות מספר שנות בצורת בשנים 2004–2009, שהובילו לקריסה סופית ולתמותה ב-2010 של חלק מהעצים (מסוג '0') (איור 3ב), וזאת בניגוד להתאוששות שאפיינה את שאר העצים (בעיקר שלא סבלו מעקה) לאחר הבצורות של 1984 ו-1996. שנות בצורת אלה ושנות הבצורת התכופות בתקופה 2000–2010 הן שנות 'תפנית', ורגישות התגובה של העצים מאפשרת לחזות את עמידות העצים ואת תגובתם בהינתן מחזוריות של שנות בצורת.
דיון
המסקנות מבוססות על תוצאות המחקר בשילוב מחקר ארוך טווח המתקיים ביער יתיר זה 15 שנים. בתנאי יובש, באזור שחון למחצה כיתיר, סיכויי שרידות והתפתחות עצי אורן ירושלים תלויים במידה רבה במאפייני בית הגידול המקומי. מתוצאות המחקר עולה כי בית גידול קרקעי, עם מיעוט אבנים וקרקע עמוקה, מעל סלע אֵם של אבן גיר, הוא בית גידול בעל רגישות גבוהה, שהעצים בו מועדים לתמותה בעקבות שנות יובש קיצוניות. מנגד, בית גידול המתאפיין בקרקע רדודה וביחס סלע/קרקע גבוה, ונמצא על בסיס סלע קירטון, הוא בית גידול מתאים יותר להישרדות עצי האורן באותם תנאים.
מכלול ההשפעות ההדדיות בין הקרקע, הסלע ושורשי העצים מורכב ובעל משמעויות רבות. סלע הקירטון הנקבובי, שנמצא בעיקר בחלקות ה'עמידות', מסתמן כמאגר אפשרי למים בעת יובש באמצעות שחרור מים אִטי בשיטת ה'ספוג'. הדבר נמצא באתרים אחרים [2], אך נדרשת בדיקה אמפירית ספציפית לתנאי יער יתיר. תופעה זו רלוונטית בעיקר בימי הקיץ, שמאופיינים בזמינות מים נמוכה, ואילו בחורף היא גורם שולי יחסית. מצד שני, היעדר כיסוי סלעי בפני השטח עשוי להגביר אידוי מפני הקרקע, והיעדר סלעים נקבוביים (קירטון) בעומק מפחית גם את כמות המים הזמינים, ועל כן, במצב כזה היעדרם יעצב בצורה שונה את פיזור מערכת השורשים ואת צפיפותה. לאור ממצאי המחקר ניתן להעריך שהימצאות סלעים מקטינה את קצב האידוי לאוויר, ומצד שני מגבירה אגירת מים בכיסי קרקע מנגר מקומי, ובכך, את כמות המים הזמינים לעצים שגדלים בבתי גידול אלה, על פי המשוואות הבאות:
(1) Pe = P (1 – (I + (Es,sAs + Es,rAr)))
כשרכיב הנגר העילי וחדירת המים מתחת לבית השורשים זניחים, וכאשר:
Pe – המים הזמינים לעץ ליחידת שטח
P – כמות המשקעים השנתית ליחידת שטח (P מ"מ בשנה)
I – אצירה (%, מהממוצע הרב-שנתי, טיפות גשם המתאדות ישירות מנוף העץ)
Es,s – התאיידות מקרקע ללא כיסוי סלעי (כלומר מהאדמה, כאחוז ממוצע מכמות המשקעים השנתית)
Es,r – ההתאדות מפני סלע שעל הקרקע [23] Es,s >>Es,r
As, Ar – הכיסוי היחסי של פני שטח סלעיים וקרקעיים בהתאמה.
על פי Raz-Yaseef [18] בתנאי יער יתיר, כ-36% מהגשם היורד מתאדה ישירות מהקרקע, בקרקע נטולת אבנים וסלעים. כאשר במקרה דנן, יחס הסלע/קרקע היה 36% ו-8% בחלקות ה'עמידות' וה'רגישות' בהתאמה, וההשפעה על כמות המים הזמינים במקרה זה משמעותית. התייחסות לאחוז האבנים בפרופיל הקרקע משנה אף היא את כמות המים האגורה ביחידת נפח אדמה ביחס לפרופיל שאין בו התייחסות לאבנים, ואת המים הזמינים לעץ, ומובילה לפיתוח משוואה נוספת:
(2)Pe* = Pe/V= Pe/(Vs Cs,W)+(Vr Cr,w))
כאשר Pe* – תכולת מים ליחידת נפח קרקע (V/V) כתלות בהרכב האדמה והאבן
Vs – נפח האדמה
Vr – נפח הסלע ביחידת נפח קרקע (Vs+Vr=V)
Cs,w – קיבול המים באדמה (%)
Cr,w – הקיבול בנפח מסוים המורכב מאבנים
Pe – ביחידות נפח (דוגמה לחישוב משוואות אלה עם נתונים אמפיריים ניתן למצוא בנספח).
מקובל לחשוב כי במרבית סוגי הקרקע יכולת אגירת המים של הקרקע גבוהה מזו של סלעים (Cr,w < Cs,w), אך בפועל, במהלך התקופה הרטובה ריכוז המים בקרקע סלעית ובעלת אחוז גבוה של אבנים, גבוה מאשר בקרקע עם אחוז אבנים נמוך. מנגד, בעונת היובש, בעת שהאדמה יבשה, סלעים אלה מספקים מים האצורים בהם, בשחרור אִטי, עקב הפרשי הפוטנציאלים בין מתח המים הנמוך שבסלע לזה הגבוה שבאדמה היבשה. עובדה זו, בשילוב שכבות הקרקע העמוקות (שורשים נצפו ביער יתיר בעומק של 6 מטרים), מאפשרת לעץ ליהנות ממקור אספקת מים אִטי אך יציב לאורך כל העונה היבשה עקב הגברת יכולת החידור של המים לעומק. עם זאת, נדרש מחקר נוסף לכימות ולהבנת הדינמיקה בזמן של זמינות המים לעץ בקרקעות שיש בהן אחוזים שונים של אבנים, ולהבנת השפעת אחוז האבנים על חדירת שורשי העצים לעומק.
מכאן, שבהנחה שבמהלך החורף מידת ספיחת המים לסלע מועטה (אך שחרורה משמעותי במהלך הקיץ כפי שנמצא במחקרים קודמים) [2], נוכחות אבן וסלע משפרת את משטר המים השנתי ליחידת נפח (V) בתווך הקרקעי, שם מצויים שורשי העצים. נוסף על כך, להגברת זמינות המים בעונה הרטובה יש משמעות ניכרת לעצים בתנאי קיצון כביתיר, מאחר שיש יותר מים זמינים כאשר תנאי הסביבה (טמפרטורה וגירעון הלחץ באדי המים [– Vapour Pressure Deficit VPD]) נוחים יותר בתקופת שיא פעילות העצים [16].
כפי שעולה מערכים שנמדדו במחקר זה, ישנה השפעה מהותית על זמינות המים לעצים באזור יובשני, ונראה שחלקה סלעית אכן מעניקה יתרון דרמטי להתפתחות ולהישרדות עצים בחלקות. מאחר שביתיר הקרקע אינה מגיעה לרטיבות של קיבול שדה במהלך החורף, עצם נוכחות הסלע בתא שטח מסוים מגבירה את כמות המים הזמינים לעץ המצוי באותו תא שטח.
תחרות על מקורות מים ומשאבים אחרים היא גורם משמעותי ביכולת הישרדות היער, ולכן צפיפות החלקה (מספר עצים לדונם), נחשבת באופן מסורתי למדד יערני משמעותי. השפעה שלילית מתונה של הצפיפות על קוטר העצים נצפית ביער (איור 3 בנספח) בעיקר בחלקות ה'עמידות', נתון המעיד על חשיבות נמוכה לתחרות על מקורות מים, על חומרי הזנה ועל אור בחלקות ה'רגישות' לעומת ה'עמידות'. התוצאות מראות שהצפיפות הנוכחית (החל באמצע שנות ה-90, כ-30 עצים לדונם) תואמת את התנאים ביתיר כיום, וחשיבותה במדיניות הממשק בהתמודדות עם תמותה הקשורה לתופעת הבצורות – משנית.
המחקר בהיבט הפיזיולוגי של העץ הבודד מצביע על כך שההבדלים בין החלקות במצב העצים ובאיכות בתי הגידול, באים לידי ביטוי גם בתאי שטח קטנים יותר, בתוך החלקות ה'עמידות'. ההבדלים המשמעותיים בין עצים שכנים, השרויים ברמות עקה שונות, בערכי פוטנציאל המים טרום שחר ובקצב הגדילה, מעידים על מצב המערכת ההידראולית והפיזיולוגית שלהם, שנגזר מתנאי תת-הקרקע שלהם. מאחר שמדידת פוטנציאל מים בטרום שחר מעידה על פוטנציאל המים בקרקע [19, 20, 21], מתברר כי עצים שכנים חשופים לתנאים שונים משמעותית בתת-הקרקע. התנאים גורמים לקצב צימוח שונה, שמתבטא גם באורך המחטים ובעובי טבעות העצים.
אם כן, מבחינת השערות המחקר ניתן לומר כי:
- לא נמצאו הבדלים גנטיים בין העצים השורדים לעצים המתים שיכולים להסביר את הכתמיות, ולכן הגורם הגנטי לא הוצג במחקר זה;
- כתמיות התמותה אכן נובעת מההבדלים הגאומורפולוגיים בין תאי שטח השונים, אך יותר בחלקה עצמה ובין עצים בודדים;
- שיפוע, מפנה, גיל וצפיפות החלקה הם גורמים משניים ולא משמעותיים בסיכויי ההישרדות של העצים;
- ישנה שונות מובהקת במבנה תת-הקרקע של החלקות השורדות לעומת החלקות ה'רגישות', דבר המוביל לשינוי במשק המים של העצים.
יערות אורנים על סף המדבר רגישים יותר לתנאי האקלים המשתנים. אי לכך, הטיפול היערני צריך להתאים לתנאי עקה ויובש קיצוניים. אבחון סימני דעיכה מוקדמת של עצים, מספר שנים לפני התמותה הסופית כפי שעולה מעובי הטבעות השנתיות (איור 3), מאפשר זיהוי מוקדם של עצים השרויים בעקה. דילול מבוקר ומושכל של עצים מסוימים, שסיכויי הישרדותם נמוכים, יכול להקל על עצים חזקים יותר להתפתח ולעמוד בשנות בצורת קשות ולהפחית במעט את התחרות שבכל זאת יש לה חשיבות בתנאי יובש. כדי להרחיב את מסקנות המחקר למערכות אקולוגיות אחרות יש צורך בהרחבת המחקר לאתרים שונים לאורך מפל המשקעים בישראל.
מסקנות
זמינות המים לעצים היא המרכיב העיקרי המשפיע על מצב היער בסף המדבר, ושרידותם תלויה בניהול ובממשק של היער, העומד והעץ הבודד. נראה, כי בגיל היער הנוכחי, בצפיפותו הנוכחית, ובמאזן הידרולוגי סגור [18], אנו מצויים על סף הצפיפות המיטבית [22]. לאור ממצאי מחקר זה, המראים כי הצפיפות היא גורם משני בהישרדות ובהתפתחות עצים ביער יתיר, דילולים ונטיעות צריכים להיערך לאחר בחינה של תנאי בית הגידול ותת-הקרקע ברזולוציה גבוהה, בהתחשב בשונות בין קבוצות עצים קטנות בבתי גידול שכנים.
שימוש במסקנות המחקר שלפניכם יאפשר זיהוי בתי גידול קרקעיים לעומת סלעיים באמצעות בחינה גאומורפולוגית פשוטה של היער, ובהתאם לתוצאותיה יתאפשר זיהוי של האזורים הרגישים לתמותה, כגון עמקים רחבים המאפיינים בתי גידול עם קרקע עמוקה, לעומת מדרונות סלעיים המאפיינים בתי גידול סלעיים.
אם כן, החלטות ממשק עתידיות צריכות להתקבל באשר לעץ וגם באשר לחלקה או לעומד. בתחום זה יש להמשיך ולפתח גם מדדים להערכת טיב תת-הקרקע המקומי שאינם תלויים בנוכחות העצים, כפי שנעשה כאן, ולשלבם בסקרי הכנה לנטיעות. יש בתוצאות מחקר זה קריאת כיוון לבחינה מחודשת של ההכנות לנטיעה מבחינת הממשק, לבחינה מחודשת של החלקה בהשוואה לעץ הבודד, ולהבנת ההטרוגניות של תא השטח מבחינת המחקר והממשק כאחד. מאחר שמחקר זה לא יכול להקיף את כל הגורמים ולהעמיק בכל אחד מהנושאים שנחקרו, יש צורך במחקר נוסף ומעמיק שיעסוק בהשפעת היחס סלע-קרקע על משק המים של העצים בבית הגידול הנבחן, באתרים שונים לאורך מפל האקלים בישראל.
לסיום, נציין כי למרות התמותה הנרחבת שבאה לאחר מספר שנות בצורת קשות, כ-95% מעצי היער שרדו, ועובדה זו מצביעה על יכולת השרידות הגבוהה מאוד של עצי אורן ירושלים, על עמידותם ועל יכולת ההתאמה שלהם לתנאי יובש.

- תמותת עצים כתוצאה משינוי האקלים היא בעיה שיערנים מתמודדים איתה בכל העולם. בארץ, יערות אורנים על ספר המדבר רגישים במיוחד לשינוי האקלים.
- במחקר נבחנו הסיבות לתמותה נרחבת של עצי אורן ירושלים ביער יתיר לאחר מספר שנות בצורת, לנוכח תצפיות של פיזור לא אחיד במרחב של העצים המתים.
- מסתבר שמאפיינים מקומיים של בית הגידול – במיוחד המסלע והקרקע – משפיעים במידה רבה על זמינות המים לשורשי העצים, ומכאן על סיכויי ההתפתחות והשרידות של עצי אורן ירושלים באזורים יובשניים למחצה, ואילו לצפיפות החלקה אין השפעה משמעותית.
- במאמר מוצעת בחינה גאומורפולוגית פשוטה של היער, המאפשרת זיהוי של האזורים הרגישים לתמותה.
- תוצאות המחקר מקדמות את ההבנה של הגורמים הקשורים לתמותת עצים ביערות, ומציעות למנהלי יערות באזורים יובשניים למחצה כלי שצריך להנחות פעולות ממשק ביער, בייחוד דילולים ונטיעות.
המערכת
נספחים
נספח 1. צילום אוויר של יער יתיר, עם שכבת ממ"ג של התמותה וחלקות המחקר
להורדהנספח 2. דוגמאות לכתמיות התמותה במהלך 2010
להורדהנספח 3. קוטר העצים, המשמש כמדד לחיוניותם, כתלות בצפיפות החלקה, בחלקות העמידות והרגישות
להורדהנספח 4. השפעת המשקעים על אורך המחטים בקטגוריות השונות והשונות באורך המחטים בשנים השונות כתלות בקטגוריה ובמשקים
להורדהטבלה 1. פירוט ערכי המשתנים אשר בהם השתמשנו לצורך הצבת הנתונים במשוואות
להורדהמקורות
- דורמן מ, סבוראי ט ופרובולוצקי א. 2012. התייבשות עצים ביערות אורן ירושלים בישראל – מבט מגובה רב. אקולוגיה וסביבה 3(3): 230–237.
- הר נ. 2008. מבנה מערכת הקרקע והסלע והדינמיקה של משק המים בבית הגידול כגורמים אקולוגיים עיקריים בתפוצת אלון התבור והאלון המצוי באזור אלונים-מנשה (עבודה לקבלת תואר דוקטור). ירושלים: האוניברסיטה העברית בירושלים.
- קק"ל. 2013. נתוני יערות קק"ל לשנת תשע"ג.
- שילר ג, אונגר י"ד וגניזי א. 2005. האם גורל העץ כתוב בטבעות השנתיות? אפיון עצים ביער כרמים בעקבות החורף השחון 1998/99. יער 7: 18–25.
- Adams H, Guardiola-Claramonte M, Barron-Gafford G, et al. 2009. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. PNAS – Proceedings of the National Academy of Sciences 106: 7063-7066.
- Allen CD, Macalady AK, Chenchouni H, et al. 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259: 660-684.
- Anderegg WRL, Berry J, and Field C. 2012. Linking definitions, mechanisms, and modeling of drought-induced tree death. Trends in Plant Science 17: 693-700.
- Anderegg WRL, Anderegg LDL, Sherman C, and Karp DS. 2012. Effects of widespread drought-Induced Aspen mortality on understory plants. Conservation Biology 26(6): 1082-1090.
- Atzmon N, Moshe Y, and Schiller G. 2004. Ecophysiological response to severe drought in Pinus halepensis Mill. trees of two provenances. Plant Ecology 171: 15-22.
- Bigler C, Gavin DG, Gunning C, and Veblen TT. 2007. Drought induces lagged tree mortality in a subalpine forest in the Rocky Mountains. Oikos 116(12): 1983-1994.
- Bonneh O. 2000. Management of planted pine forests in Israel: Past, present and future. In: Neeman G and Trabaud L (Eds). Ecology, biogeography and management of Pinus halepensis and P. brutia forest ecosystems in the Mediterranean basin. Leiden (The Netherlands): Backhuys Publishers.
- Breshears DD, Myers OB, Meyer CW, et al. 2009. Tree die-off in response to global change type drought: Mortality insights from a decade of plant water potential measurements. Frontiers in Ecology and the Environment 7(4): 185-189
- Calamassi RG, Della Rocca M, Falusi E, et al. 2001. Resistance to water stress in seedlings of eight European provenances of Pinus halepensis Mill. Annals of Forest Science 58(6): 663-672
- Grünzweig JM, Gelfand I, Fried Y, and Yakir D. 2007. Biogeochemical factors contributing to enhanced carbon storage following afforestation of a semi-arid shrubland. Biogeosciences 4: 891-904
- Hinckley TM, Lassoie JP, and Running SW. 1978. Temporal and spatial variations in the water status of forest trees. Forest Science 24: a0001-z0001
- Maseyk K, Lin T, Rotenberg E, et al. 2008. Physiology-phenology interactions in a productive semi-arid pine forest. The New Phytologist 178: 603-616
- McDowell NG. 2011. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiology 155: 1051-1059.
- Raz-Yaseef, N, Rotenberg E, and Yakir D. 2010. Effects of spatial variations in soil evaporation caused by tree shading on water flux partitioning in a semi-arid pine forest. Agricultural and Forest Meteorology 150: 454-462.
- Reich P and Hinckley T. 1989. Influence of pre-dawn water potential and soil-to-leaf hydraulic conductance on maximum daily leaf diffusive conductance in two oak species. Functional Ecology 3(6): 719-726.
- Ritchie GA and Hinckley TM. 1975. The pressure chamber as an instrument for ecological research. Advances in Ecological Research 9: 165-254.
- Sala OE, Lauenroth W, Parton W, and Trlica M. 1981. Water status of soil and vegetation in a shortgrass steppe. Oecologia 48: 327-331.
- Ungar ED, Rotenberg E, Raz-Yaseef N, et al. 2013. Transpiration and annual water balance of Aleppo pine in a semiarid region: Implications for forest management. Forest Ecology and Management 298: 39-51.
- Shi Z, Wang Y, Yu P, et al. 2008. Effect of rock fragments on the percolation and evaporation of forest soil in Liupan Mountains, China. Acta Ecologica Sinica 28(12): 6090-6098.



 מהדורה מודפסת
מהדורה מודפסת







