
חזית המחקר
רעייה ובצורת בספר המדבר – מגמות ארוכות טווח בכיסוי הצומח המעוצה
יוג׳ין קנדל המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקני
הילרי פוט הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושלים
רפי יונתן המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקני
דני ברקאי המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקני
עזרא בן משה המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקני
חגית ברעם המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקני
חיים קיגל הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושלים

קיימת השפעה הדדית בין כמות השיחים במדבר וסוגיהם לבין רעיית הצאן. הבצורת עשויה להוביל לירידה בשטח המכוסה על-ידי שיחים | צילום: ניב דה מלאך
ניב דה מלאך
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםיוג׳ין קנדל
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניהילרי פוט
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםרפי יונתן
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקנידני ברקאי
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניעזרא בן משה
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניחגית ברעם
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניחיים קיגל
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםניב דה מלאך
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםיוג׳ין קנדל
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניהילרי פוט
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםרפי יונתן
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקנידני ברקאי
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניעזרא בן משה
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניחגית ברעם
המחלקה לגידולי שדה ומשאבי טבע, המכון למדעי הצמח, ִמנהל המחקר החקלאי – מרכז וולקניחיים קיגל
הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושליםרעייה היא שימוש קרקע נפוץ ביותר במערכות אקולוגיות צחיחות למחצה שהגורם המגביל בהן לפעילות ביולוגית הוא מים. שיחים הם מרכיב חשוב של מערכות אלה, הן בשל השפעתם הרבה על תהליכים המתרחשים במערכת האקולוגית הן בשל השפעתם על ערך השטח כמרעה. מטרת המחקר הייתה לבדוק את ההשפעה של רעיית צאן, בצורת והשילוב ביניהן על הצומח המעוצה במערכת צחיחה למחצה. המחקר נערך באזור גבעות גורל ליד היישוב להבים, בספר המדבר בצפון הנגב בשנים 1996–2007. בחנו את הכיסוי של מיני השיחים הנפוצים (סירה קוצנית, מִתנן שעיר ונואית קוצנית) בארבעה בתי גידול: מפנה צפוני, מפנה דרומי, ראש גבעה וכתפי ואדי. כדי לבדוק את השפעת הרעייה על השיחים הוקמו גדוּרות להגנה מרעייה. הכיסוי היחסי של השיחים נמדד בשיטת point transect. המודל הסטטיסטי כלל ניתוח שונות תלת-גורמי לבדיקת מובהקות השפעות הרעייה, בית הגידול והשנה (כמשתנים בדידים). כמו כן, נבחנה השפעת שתי שנות בצורת רצופות (1999 ו-2000) על מגמות ארוכות טווח בכיסוי השיחים.
תוצאות המחקר מראות כי כיסוי השיחים בחלקות ללא רעייה במשך 11 שנים היה גדול יותר מאשר בחלקות תחת רעייה רציפה (21% לעומת 16%), אם כי השפעת הרעייה הייתה שונה בין המינים ובין בתי הגידול השונים. לבית הגידול הייתה השפעה גדולה ומובהקת על הכיסוי המעוצה. בעוד שבמפנה הצפוני שלטה סירה קוצנית, בשאר בתי הגידול לא היה מין אחד דומיננטי. לא נמצאו אינטראקציות בין הרעייה לבין השנים, למרות השונות הגדולה במשקעים בין השנים. דגמי התגובה של השיחים לאירוע הבצורת היו שונים בין המינים ובין בתי הגידול – בעוד שרמת הכיסוי של הנואית הקוצנית עלתה באופן זמני בהשפעת הבצורת, רמת הכיסוי של הסירה הקוצנית והמתנן השעיר ירדה בעקבות הבצורת, עם התאוששות מוגבלת עבור סירה קוצנית. תוצאות המחקר מעלות חשש באשר ליציבות המערכת האקולוגית בספר המדבר בצפון הנגב, וזאת בשל שינוי האקלים הצפוי להתרחש באזור. אירועי בצורת תכופים עלולים לגרום לתמותה מְבַדלת (differential) של מיני שיחים, להקטנת צפיפות כלל השיחים ולשינוי בהרכב כיסוי השיחים.
על קצה המזלג
- אזור ספר המדבר מתאפיין בשטחי מרעה נרחבים ובמיעוט משקעים המגביל את פוריות המערכת האקולוגית.
- לשיחים תפקיד מרכזי בספר המדבר בגלל מרכזיותם במערכת האקולוגית ובגלל חשיבותם לרעיית צאן.
- המחקר בדק את ההשפעה של רעייה ושל שנות בצורת על כיסוי השיחים הנפוצים בשטח צחיח למחצה.
- נמצא כי לרעייה ולבצורת יש השפעה על כיסוי השטח על-ידי שיחים, התלויה במין השיח ובאופי בית הגידול, וכי השפעת הבצורת משמעותית יותר.
- תוצאות המחקר מעלות חשש ששינוי האקלים הצפוי עלול להביא לתמותה של מיני שיחים מסוימים ולירידה בכיסוי השטח הכללי על-ידי שיחים.
הלכה למעשה
מבוא
רעייה היא שימוש הקרקע הנפוץ ביותר בעולם [6], בפרט באזורים צחיחים למחצה [15]. ברם, טיב השפעת הרעייה על מערכות אקולוגיות באזורים צחיחים למחצה הוא נושא שנוי במחלוקת. יש הטוענים שרעייה על-ידי אוכלי צמחים מבויתים היא אחד הגורמים העיקריים למדבור ולהתדרדרות המערכת האקולוגית באזורים הללו [19]. לעומתם, טוענים אחרים שבאזורים כמו המזרח התיכון, הנמצאים כבר אלפי שנים תחת רעייה, השפעתה של הרעייה על המערכת האקולוגית איננה שלילית בהכרח [14].
המחלוקת הזו מקבלת משנה תוקף בעידן של שינוי אקלים שיש בו חשש לפגיעה בתפקוד מגוון רחב של מערכות אקולוגיות [3, 24]. שינוי במשטר הגשמים עלול לשנות את תפקוד המערכת האקולוגית באזורים צחיחים למחצה מפני שזמינות מים היא הגורם העיקרי המשפיע על תהליכים ביולוגיים באזורים הללו [11]. בשני העשורים האחרונים העוצמה והתכיפות של בצורות באזורים רבים בעולם גדלו מאוד [4], מגמה הצפויה להתגבר בעתיד, במיוחד באזורים צחיחים למחצה [16].
שינוי אקלים ושינוי בשימושי קרקע נחשבים כאיומים הגדולים ביותר על תפקוד מערכות אקולוגיות. אף על פי שאיומים אלה עלולים לפעול באותו זמן, לרוב הם נחקרו בנפרד, למרות הבעייתיות שבהפרדה זו [5]. על כן, מחקר ארוך טווח שבוחן שינויים החלים בשטחים הנתונים לרעייה לעומת שטחים ללא רעייה, עשוי לתרום להבנת הקשרים בין ההשפעות של שינוי אקלים והשינוי בשימושי קרקע, במיוחד באזורים צחיחים וצחיחים למחצה. באזורים אלה שיחים עשויים להיות סמן טוב לתפקוד המערכת האקולוגית בגלל השפעתם הרבה על תהליכים המתרחשים בה, כגון שטפי פחמן וחנקן [27], פיזור מי נגר [25] ונשימת הקרקע [23], וכן גם על השפע והפיזור של שאר האורגניזמים [21].
מטרת המחקר הנוכחי הייתה לבחון את מידת העמידות של השיחים לרעייה, לבצורת ולשילוב ביניהן. ניצלנו את השנים 1999 ו-2000 שהתאפיינו בבצורת קשה, כדי לבחון דגמי תגובה של כיסוי הצומח המעוצה לבצורת בסקלת זמן של עד שבע שנים מהבצורת. הנחנו שבעקבות הבצורת יכולים להתרחש חמישה דגמי תגובה תאורטיים של שינוי בכיסוי השיחים: א) חוסר השפעה; ב) השפעה שלילית – ירידה בכיסוי ללא התאוששות; ג) השפעה חיובית – עלייה בכיסוי והתייצבות; ד) פגיעה והתאוששות – ירידה בכיסוי ועלייה חזרה למצבו ההתחלתי; ה) עידוד זמני – עלייה בכיסוי וירידה למצבו ההתחלתי. להבדיל מצומח עשבוני, קשה מאוד למדוד ביומסה של שיחים. מסיבה זו מקובל להעריך את השינויים החלים במרכיב השיחים בצומח על-ידי אומדן הכיסוי שלהם והשינויים החלים בו במשך הזמן ובתגובה לתנאים סביבתיים, כולל רעייה.
חומרים ושיטות
שטח המחקר
הניסוי התבצע בלהבים, אתר למחקר ארוך טווח (LTER) בספר המדבר בנגב הצפוני (31º 20’ N, 34º 45’ E). האזור נתון לרעיית כבשים ועִזים מזה שנים רבות [1]. כיום רועה בחווה עדר של 600 כבשים ו-200 עִזים כ-7 שעות ביום. בלילה לן העדר במכלאה. לחץ הרעייה משתנה בין אזורים שונים בחווה, אך עומד בממוצע על 10 דונם לראש. העדר מקבל תוספת הזנה במהלך חודשי הקיץ והסתיו. כמות המשקעים הממוצעת (± סטיית תקן) היא 298±90 מ"מ לעונה, עם שינויים גדולים מאוד לאורך שנות הניסוי – מעונת 1995/96 ועד 2006/07 (להלן שנים 1996 עד 2007) (איור 1). העונות 1998/99 ו–1999/2000 (להלן שנים 1999 ו-2000) התאפיינו בבצורת קשה, עם 140 ו-198 מ"מ גשם לשנה, בהתאמה.
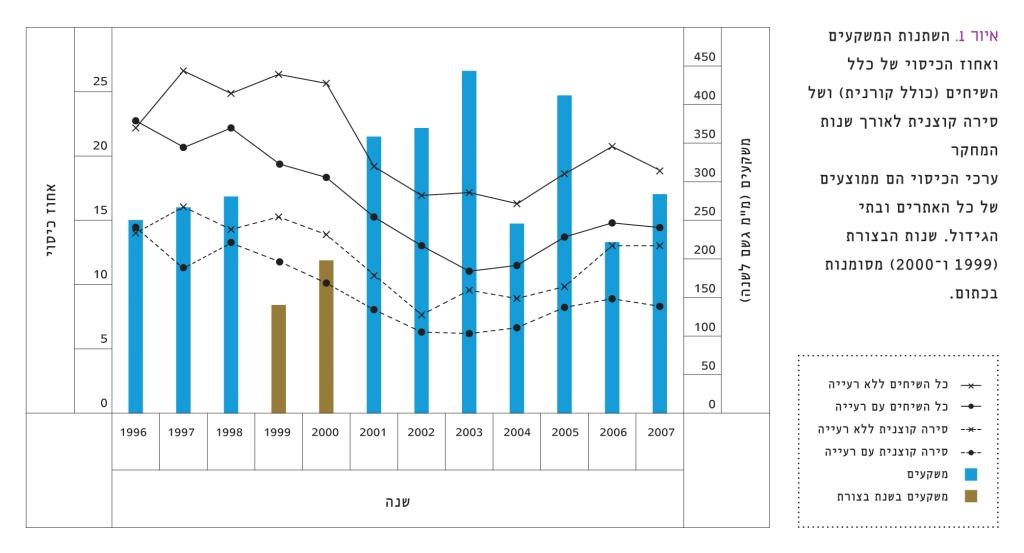
איור 1
השתנות המשקעים ואחוז הכיסוי של כלל השיחים (כולל קורנית) ושל סירה קוצנית לאורך שנות המחקר
ערכי הכיסוי הם ממוצעים של כל האתרים ובתי הגידול. שנות הבצורת (1999 ו-2000) מסומנות בכתום.
השיח הנפוץ ביותר מבין השיחים בשטח המחקר הוא סירה קוצנית (Sarcopoterium spinosum), בן שיח שבולט במיוחד במפנים הצפוניים [22]. בני שיח נוספים בעלי תפוצה רחבה באזור, שנפוצים יותר במפנים הדרומיים, הם מִתנן שעיר (Thymelaea hirsuta), נואית קוצנית (Noaea mucronata) וקורנית מקורקפת Coridothymus capitatus) [1]).
מבנה חלקות המחקר וחתכי צומח
הניסוי נערך בשלושה אגני ניקוז שונים (אתרים) בשטח חוות המחקר. בכל אתר מיוצגים ארבעה בתי גידול שונים: מפנה צפוני, מפנה דרומי, ראש גבעה וכתף ואדי. בתי הגידול נבדלים ביניהם בחשיפה לקרינת השמש, בטמפרטורה, בכמות הנגר שהם מקבלים או מאבדים, וגם בסלעיות ובעומק הקרקע. בכל צירוף של אתר ובית גידול הוקמו 4 חלקות (סך הכול 48 חלקות). כל חלקה הייתה מורכבת משתי תת-חלקות: גדורה בגודל של 10×10 מטר ולצִדה שטח מסומן בגודל דומה, שנמצא מחוץ לגדורה ופתוח לרעייה. הגדורות הוקמו בין השנים 1994–1996. למרות השוני בתאריך הקמתן לא נעשתה הבחנה בין הגדורות, מתוך הנחה שהבדל של שנה או שנתיים בגידור הוא קטן יחסית במחקר ארוך טווח. מתוך 48 חלקות מקוריות נבחרו 36 חלקות שההפרש בכיסוי השיחים בין תתי-החלקות בהן בשנת 1996 (תחילת הניסוי) לא עלה על 5% מסך הכיסוי הכללי. הבדלים גדולים יותר עשויים להעיד על שוני בתשתית החלקה (סלעיות, עומק וטיב הקרקע), ולכן נבחרו לניתוח חלקות שתנאי ההתחלה בהן היו דומים ככל האפשר. ניטור כיסוי השיחים נערך כל שנה בקיץ, בין השנים 1996 ו-2007. נתוני הכיסוי בחלקות התקבלו מחתכים קבועים באורך של 10 מטר, מסומנים ביתדות, בשיטת point transect. כל 10 ס"מ לאורך החתכים בדקנו אם קיים כיסוי מעוצה, ואם כן – לאיזה מין שיח הוא שייך. אחוז הכיסוי של כל מין מוצג ככיסוי באחוזים מכלל אורך החתך. בכל חלקה היו שני חתכים: אחד בתוך הגדורה (שטח ללא רעייה) ואחד מחוץ לה (שטח תחת רעייה).
ניתוח סטטיסטי
הניתוח הסטטיסטי נעשה באמצעות תכנת 9.1 SAS. המשתנה התלוי היה הכיסוי היחסי של צומח מעוצה לאחר טרנספורמציה של arcsin שהתאימה את נתוני הכיסוי להתפלגות נורמלית. נבדקו מודלים נפרדים למינים סירה קוצנית, מתנן שעיר ונואית קוצנית, ולכיסוי המעוצה הכללי (כולל קורנית מקורקפת). הניתוח הראשון היה ניתוח שונות רב-גורמי עם מדידות חוזרות (ANOVA with repeated measures). לשם כך הגדרנו "שנה" כגורם בדיד שרירותי שמגלם בתוכו את מכלול התנאים הסביבתיים שמאפיינים שנה ושיכולים להסיט את כלל המדידות של שנה מסוימת בכיוון מסוים. יחידת המדידה הייתה חלקה, מקננת (nesting) בתוך בית גידול ואתר. כל חלקה חולקה לשתי תתי-חלקות: עם רעייה ובלעדיה. ההשוואות בין בתי הגידול נעשו באמצעות מבחן α=0.05) Tukey). השפעת הרעייה נבחנה במבנה של טיפולים מזוּוגים (matched pairs) בין חתכים בתוך הגדורה ומחוץ לגדורה.
קורנית מקורקפת הראתה תבנית פיזור חריג: היא נמצאה בשכיחות גבוהה במפנה הדרומי של אתר אחד, ובשאר האתרים ובתי הגידול השכיחות שלה הייתה נמוכה מאוד. לאור השוני בתבנית הפיזור ועקב חוסר ההומוגניות של השונות, מודל כללי של ניתוח שונות לא התאים, ולכן היא לא נבחנה במסגרת הניתוח הסטטיסטי שצוין לעיל.
בניתוח השני נבחנה התגובה של כיסוי השיחים לגשם. נעשתה רגרסיה לינֵארית בין כמות המשקעים השנתית (בפועל, מאוקטובר עד מאי) לבין השינוי –היחסי או המוחלט – בכיסוי השיחים הכללי, בתנאי רעייה וללא רעייה.
בניתוח השלישי סיווגנו את דגמי השינוי בכיסוי שהתקבלו בתגובה לבצורת לפי הדגמים השונים שתוארו לעיל. לשם כך השווינו את הכיסוי בכל אחת מהשנים מתחילת הבצורת (1999) ועד סוף הניסוי (2007) מול ממוצע הכיסוי בשנים 1996–1998, שקדמו לבצורת ושהתאפיינו בכמות משקעים ממוצעת. השוואה כזאת נעשתה בנפרד עבור כל המינים בכל בתי הגידול, ועבור הכיסוי המעוצה הכללי בכל בתי הגידול. השתמשנו במבחן (Dunnett (α=0.05 שמיועד להשוואת ערכים רבים לערך אחד. מקרים שלא הייתה בהם מגמה עקבית הוגדרו כ"חוסר השפעה". מקרים שהכיסוי בהם אחרי השנים 1999–2002 היה נמוך באופן מובהק מזה שבשנים 1996–1998 ונשאר נמוך מאז ועד סוף תקופת הניסוי הוגדרו כ"השפעה שלילית" (גם אם התקבלה מגמה של התאוששות קלה). מקרים שהכיסוי בהם בשנים 1999–2002 היה גבוה באופן מובהק מהכיסוי בשנים 1996–1998 ונשאר גבוה גם לאחר מכן הוגדרו כ"השפעה חיובית". מקרים שהכיסוי בהם בשנים 1999–2002 היה נמוך באופן מובהק מזה שבשנים 1996–1998 אך חזר בסוף הניסוי לרמת הכיסוי ההתחלתית הוגדרו כ"פגיעה והתאוששות". מקרים שהכיסוי בהם בשנים 1999–2002 היה גבוה באופן מובהק מזה שבשנים 1996–1998 אך חזר בסוף הניסוי לרמת הכיסוי ההתחלתית הוגדרו כ"עידוד זמני".
תוצאות
מגמות כלליות בכיסוי השיחים
נמצאו הבדלים מובהקים בין בתי הגידול השונים (P<0.0001) ובין השנים (P<0.0001) עבור אחוז הכיסוי הכללי של השיחים וכן עבור אחוז הכיסוי של שלושת מיני השיחים העיקריים בנפרד. בניתוח הכיסוי של כלל השיחים נמצא שהשפעת הרעייה בכל האתרים ובתי הגידול הייתה מובהקת (P<0.0001), אולם היה הבדל גדול בין המינים בתגובה לרעייה בבתי הגידול השונים (טבלה 1). בכל הצירופים של מין ובית גידול שהשפעת הרעייה בהם הייתה מובהקת, כיסוי השיחים תחת רעייה היה נמוך יותר מאשר ללא רעייה: הכיסוי המעוצה הכללי היה 16.5±0.7% לעומת 21.3±0.8% בהתאמה, כיסוי הסירה הקוצנית היה 9.5±0.6% לעומת 12.2±0.7% בהתאמה, וכיסוי הנואית הקוצנית היה 1.8±0.2% לעומת 4.1±0.3% בהתאמה (בממוצע רב-שנתי של כל בתי הגידול ביחד). אצל המתנן השעיר לא היה הבדל מובהק בין הכיסוי ללא רעייה ותחת רעייה, והוא עמד בשניהם על 2.0±0.2%.

טבלה 1
השפעה של רעייה, בית גידול, שנה והאינטראקציות ביניהם על כיסוי השיחים על פי ניתוח שונות (ANOVA)
שנה היא גורם בדיד שרירותי שמגלם בתוכו את מכלול התנאים הסביבתיים שמאפיינים שנה. נבדקו שלושה אתרים שונים, ארבעה בתי גידול בכל אתר. DF – מספר דרגות חופש, F – ערך מבחן P ,F – הסתברות.
השפעת הרעייה הייתה דומה בשנים השונות עבור הכיסוי המעוצה הכללי, ועבור שלושת מיני השיחים בנפרד (האינטראקציה בין הרעייה לשנה לא נמצאה מובהקת). לעומת זאת, נמצאו אינטראקציות מובהקות בין בית גידול לשנה עבור סירה קוצנית(P<0.0001) ומתנן שעיר (P=0.042) בכל בתי הגידול למעט ראש הגבעה, אך לא עבור נואית קוצנית וכיסוי השיחים הכללי. כמו כן נמצאו אינטראקציות מובהקות בין בית הגידול לרעייה עבור סירה קוצנית (P<0.0001), מתנן שעיר (P=0.009) והכיסוי המעוצה הכללי (P=0.001), אך לא עבור נואית קוצנית. בראש הגבעה, בניגוד לשאר בתי הגידול, לא נמצאה השפעה מובהקת לרעייה עבור סירה קוצנית והכיסוי המעוצה הכללי. במפנה הצפוני, ובניגוד לשאר בתי הגידול שהשפעת הרעייה לא הייתה מובהקת בהם, כיסוי המתנן השעיר היה גבוה יותר תחת רעייה (1.1%) מאשר ללא רעייה (0.6%, P<0.01).
תגובה לגשם
לא נמצא קשר לינֵארי מובהק בין כמות המשקעים השנתית לבין השינוי המוחלט או היחסי בכיסוי השיחים, עם רעייה או בלעדיה (ערך P הקטן ביותר בין ארבעה ניתוחי רגרסיה אלה היה 0.49).
דפוסי תגובה לבצורת
בניתוח דפוסי תגובה של כיסוי השיחים שהתקבלו לאחר הבצורת בשנים 1999–2000 אוחדו נתוני הכיסוי של תתי-החלקות עם רעייה ובלעדיה, מכיוון שלא נמצאו אינטראקציות מובהקות בין רעייה לשנה (טבלה 1). בבחינת המודל של השפעת הבצורת נמצא שדגם השינוי בכיסוי השיחים היה שונה בין מיני השיחים. כמו כן, עבור אותם מיני שיחים התקבלו דגמי תגובה שונים לבצורת בבתי הגידול השונים (איור 2).
כיסוי השיחים של הסירה הקוצנית ירד בצורה מובהקת בעקבות הבצורת (P<0.05) בכל בתי הגידול בהשוואה לכיסוי ההתחלתי הממוצע בשנים 1996–1998, פרט לראש גבעה. דגמי התגובה של הסירה הקוצנית לבצורת היו כדלהלן: השפעה שלילית בכתפי ואדי, השפעה שלילית במפנה הצפוני (אם כי יש סימני התאוששות), פגיעה והתאוששות במפנה הדרומי וחוסר השפעה בראש גבעה. דגמי תגובה דומים התקבלו בכיסוי המעוצה הכללי בבתי הגידול השונים, כתוצאה מהעובדה שהסירה הקוצנית היא השיח הדומיננטי בכל בתי הגידול.
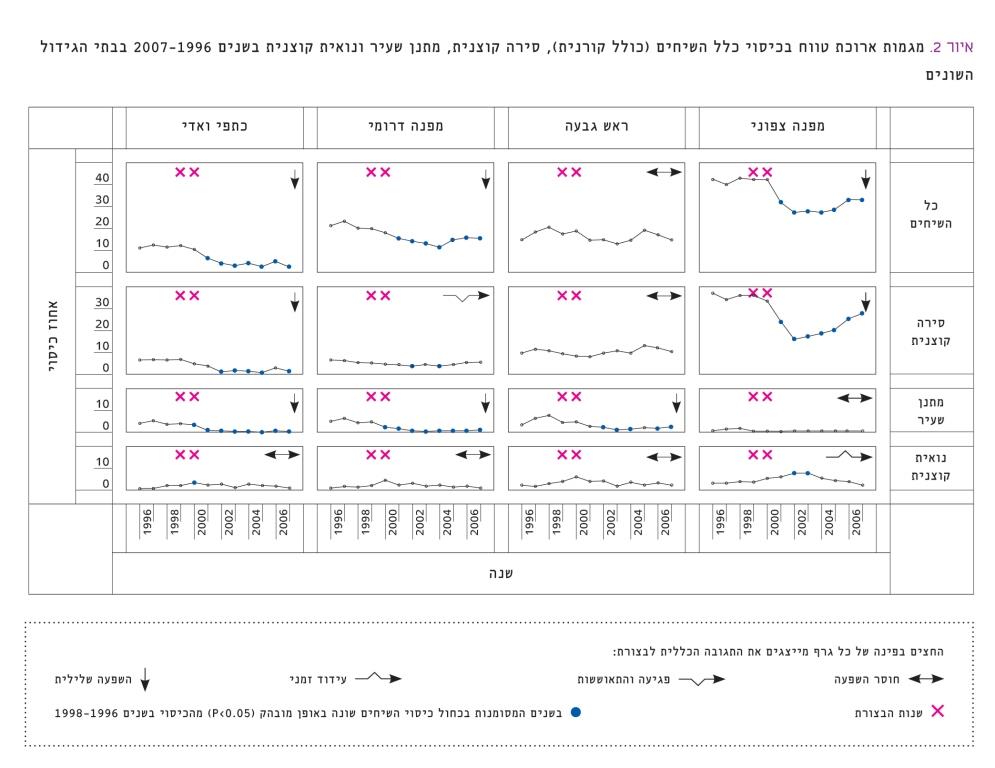
איור 2
מגמות ארוכת טווח בכיסוי כלל השיחים (כולל קורנית), סירה קוצנית, מתנן שעיר ונואית קוצנית בשנים 1996–2007 בבתי הגידול השונים
הפגיעה החמורה ביותר עקב הבצורת הייתה במתנן שעיר. אצל מין זה התקבל דגם תגובה של השפעה שלילית בשלושה מבתי הגידול, ובשניים מהם חלה ירידה מובהקת בכיסוי כבר בשנת הבצורת השנייה. במפנה הצפוני התקבל דגם תגובה של חוסר השפעה, אם כי הכיסוי ההתחלתי של המתנן השעיר היה פחות מ-1%. אחוז הכיסוי של הנואית הקוצנית הראה מגמה של עלייה זמנית בכל בתי הגידול, ומגמה זו הייתה מובהקת במפנה הצפוני. מבין בתי הגידול, כתפי הוואדי והמפנה הדרומי הראו את הפגיעה החמורה ביותר בכיסוי המעוצה הכללי – ירידה ללא התאוששות, ואילו במפנה הצפוני הייתה מגמת עלייה באחוז הכיסוי בשנות המחקר האחרונות, אך אחוז הכיסוי לא חזר למצב שהיה בו לפני הבצורת (איור 2).
התגובה של אחוז הכיסוי לבצורת התאפיינה באיחור (lag) של שנה לאחר סיום הבצורת עבור המעוצה הכללי והסירה הקוצנית, ותגובה מיידית בשנה השנייה לבצורת (2000) עבור נואית קוצנית ומתנן שעיר.
דיון
ממצאי המחקר מראים שגם לרעייה וגם לבצורת ישנה השפעה על אחוז הכיסוי של הצומח המעוצה, אך ההשפעה תלויה במין הצמח ובבית הגידול. השפעת הבצורת על אחוז הכיסוי הייתה גדולה יותר מאשר ההשפעה של הרעייה. ברם, לא נמצאה אינטראקציה בין השפעת הרעייה להשפעת השנה, למרות ההבדלים הגדולים במשקעים בין השנים.
השפעת הרעייה
הפסקת הרעייה גרמה לעלייה של 4% בכיסוי המעוצה הכללי, אך היה הבדל בהשפעתה על מיני שיחים שונים. נואית קוצנית הושפעה יותר מאשר סירה קוצנית ואילו מתנן שעיר לא הושפע כלל מהפסקת הרעייה. נראה כי ההגנות הצורניות (כלומר הקוצניוּת) של הסירה הקוצנית והנואית הקוצנית יעילות פחות מההגנות הכימיות של המתנן השעיר, שהוא צמח רעיל [17].
השפעת הבצורת
התוצאות מראות שבצורת היא אכן גורם חשוב בדינמיקה של הצומח המעוצה. כיסוי השיחים הכללי ירד ברוב בתי הגידול בסקלת הזמן של הניסוי. התאוששות מהבצורת במשך שבע השנים שלאחריה, נמצאה רק אצל הסירה הקוצנית במפנה הדרומי ובאופן חלקי במפנה הצפוני.
ממצא מעניין נוסף הוא שהירידה בכיסוי הסירה הקוצנית בעקבות הבצורת התחילה שנה לאחר סיום הבצורת ונמשכה מספר שנים לאחר מכן, למרות ששנים אלו היו גשומות מהממוצע (איור 1). לעומת זאת, הירידה בכיסוי המתנן השעיר והעלייה בכיסוי הנואית הקוצנית החלו כבר בשנה השנייה לבצורת. הבדלים אלה בין השיחים בעיתוי הירידה בכיסוי וביכולת ההתאוששות לאחר הבצורת הם כנראה הסיבה העיקרית להיעדר קשר מובהק בין כמות המשקעים לשינוי בכיסוי הכללי. התופעה של תגובה מאוחרת שנמצאה אצל הסירה הקוצנית ניתנת להסבר באמצעות השערתו של Manion [7]. לדבריו, ייתכן שלאחר עקה חמורה יכולים עצים להמשיך לדעוך במשך שנים לפני שהם מתים. הסיבה לדעיכה יכולה להיות פיזיולוגית (למשל, פגיעה במוליכות צינורות ההובלה), או היחלשות הגנות הצמחים ורגישוּת גדולה יותר למזיקים. ההשערה הזו קיבלה אישוש אמפירי במחקרים על תמותה של עצים [13, 18], אך ככל הידוע לנו לא נמצאה עד כה במחקרים על שיחים.
מיני השיחים הראו דפוסי השתנות שונים של הכיסוי בתגובה לבצורת בבתי גידול שונים. נראה שהתרחשות של מצבי תגובה שונים תלויה בשני גורמים עיקריים:
א. מין השיח – ישנם דפוסי התנהגות שחוזרים על עצמם בבתי גידול שונים בהתאם למין השיח. בעוד שהמתנן השעיר הראה ירידה בכיסוי בכל בתי הגידול, הסירה הקוצנית הראתה התאוששות בחלק מבתי הגידול והנואית הקוצנית הראתה אפילו מגמה של עלייה זמנית בכיסוי, שהייתה מובהקת בחלק מבתי הגידול. הבדלים בין המינים במאפיינים צורניים ופיזיולוגיים המשפיעים על משק המים של הצמח יכולים להסביר את השוני בתגובת מיני השיחים לבצורת [8], אולם במחקר זה לא נבדקו התכונות הללו.
ב. בית הגידול – שיחים בבתי גידול שונים הראו תגובה שונה לבצורת. המפנה הצפוני וכתפי הוואדי התאפיינו בירידה הגבוהה ביותר בכיסוי השיחים. בעוד שבמפנה הצפוני הייתה מגמה של התאוששות (בעיקר בכיסוי הסירה הקוצנית), בכתפי הוואדי – בית הגידול היצרני ביותר עבור העשבוניים [12] – לא נראתה התאוששות. באזורים צחיחים למחצה יצרנות גבוהה של בתי גידול היא לרוב תוצאה של פיזור מחדש (redistribution) של מים [26], ולכן ניתן להניח שכתפי הוואדי מקבלות תוספת נגר בשנים גשומות. אנו משערים שהשונות הגדולה בתוספת הנגר בין השנים היא הגורם הראשי לפגיעה החמורה בכתפי הוואדי בשנות בצורת. ברם, כדי להבין את ההידרולוגיה השונה בין בתי הגידול יש צורך במחקרים נוספים.
כמו כן, ניתן להניח שחוסר ההתאוששות של הסירה הקוצנית בכתפי הוואדי בשנים הגשומות שלאחר הבצורת קשור לעיכוב בהתבססות של פרטים צעירים בעקבות תחרות עם עשבוניים [20]. לעומת זאת, ייתכן שבמפנה הצפוני שנמצא בו הכיסוי הגבוה ביותר של סירה קוצנית, פגיעת הבצורת בכיסוי נובעת מהחמרת עקת היובש בנקודות שאגירת המים בתשתית מוגבלת בהן, אך מתאפשרת התאוששת בשנים גשומות יותר בגלל מקור זרעים גדול יותר ומיעוט תחרות עם הצומח העשבוני ביחס לכתפי הוואדי.
רעייה ובצורת
השערה מקובלת גורסת שיש מתאם בין עמידות לרעייה ועמידות ליובש, בין היתר בשל העובדה ששתי העמידויות נובעות מתכונות דומות של התחדשות מהירה, למשל מרקמות יוצרות (מֵריסְטֵמות) בבסיס הצמח [9]. השערה חלופית מציעה שישנן תכונות שמקנות עמידות רק נגד רעייה או רק נגד בצורת [2]. התוצאות שלנו מחזקות את ההשערה השנייה, מפני שמתנן שעיר הראה בניסוי עמידות לרעייה אך לא לבצורת, ונואית קוצנית הראתה עמידות לבצורת אך לא לרעייה.
למרות ההנחה שלרעייה יש השפעה שלילית וחריפה יותר על הצומח בשנות בצורת בשל לחץ רעייה כבד יותר על הצמחים (ולכן מומלץ להקטין אותה בשנות בצורת) [10], ולמרות ההבדלים שנמצאו בתגובה של השיחים השונים לרעייה ולבצורת, לא נמצאו השפעות גומלין בין הרעייה לבין השנים עבור כיסוי כלל השיחים. כנראה שהסיבה לכך היא שהרכב החברה המעוצה לא השתנה בצורה מהותית בעקבות הרעייה, והסירה הקוצנית נשארה המין הדומיננטי בו.
לסיכום, תוצאות המחקר מעלות חשש באשר ליציבות המערכת האקולוגית בספר המדבר בצפון הנגב, וזאת בשל שינוי האקלים הצפוי להתרחש באזור. על פי המודלים לחיזוי שינוי אקלים במזרח הים התיכון יאופיין השינוי הזה בהקטנה של כמות המשקעים ובהגדלה של תכיפות הבצורות [4, 16]. אירועי בצורת תכופים עלולים לגרום לתמותה מְבַדלת של מיני שיחים, להקטנת צפיפות כלל השיחים ולשינוי בהרכב כיסוי השיחים.
תודות
תודה לשמחה לב-ידון על דיונים בתוצאות המחקר. תודה לקרן המדען הראשי של משרד החקלאות ופיתוח הכפר, להנהלת ענף המרעה, ולקרן קיימת לישראל על מימון המחקר.
- אזור ספר המדבר מתאפיין בשטחי מרעה נרחבים ובמיעוט משקעים המגביל את פוריות המערכת האקולוגית.
- לשיחים תפקיד מרכזי בספר המדבר בגלל מרכזיותם במערכת האקולוגית ובגלל חשיבותם לרעיית צאן.
- המחקר בדק את ההשפעה של רעייה ושל שנות בצורת על כיסוי השיחים הנפוצים בשטח צחיח למחצה.
- נמצא כי לרעייה ולבצורת יש השפעה על כיסוי השטח על-ידי שיחים, התלויה במין השיח ובאופי בית הגידול, וכי השפעת הבצורת משמעותית יותר.
- תוצאות המחקר מעלות חשש ששינוי האקלים הצפוי עלול להביא לתמותה של מיני שיחים מסוימים ולירידה בכיסוי השטח הכללי על-ידי שיחים.
מקורות
- פרבולוצקי א ולנדאו י. 1988. שיפור ופיתוח ענף הצאן במגזר הבדווי בנגב הצפוני: חוות ההדגמה לבדווים – להבים. דו"ח מקצועי 1988-1982, מִנהל המחקר החקלאי, בית-דגן.
- Cingolani AM, Noy-Meir I, an d Díaz S. 2005. Grazing effects on rangeland diversity: A synthesis of contemporary models. Ecological Applications 15: 757-773.
- Cramer W, Bondeau A, Woodward FI, Prentice IC, Betts RA, Brovkin V, Cox PM, Fisher V, Foley JA, Friend AD, Kucharik C, Lomas MR, Ramankutty N, Sitch S, Smith B, White A, and Young-Molling C. 2001. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Global Change Biology 7: 357-373.
- Dai A. 2011. Drought under global warming: A review. Wiley Interdisciplinary Reviews: Climate Change 2: 45-65.
- de Chazal J and Rounsevell MDA. 2009. Land-use and climate change within assessments of biodiversity change: A review. Global Environmental Change 19: 306-315.
- Lund HG. 2007. Accounting for the world's rangelands. Rangelands 29: 3-10.
- Manion PD. 1981. Tree disease concepts. NJ: Prentice Hall.
- McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, and Yepez EA. 2008. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist 178: 719-739.
- Milchunas DG, Sala OE, and Lauenroth WK. 1988. A generalized model of effects of grazing by large herbivores on grassland community structure. American Naturalist 132: 87-106.
- Milton SJ, Dean WRJ, du Plessis MA, and Siegfried WR. 1994. A conceptual model of arid rangeland degradation. BioScience 44: 70-76.
- Noy-Meir I. 1973. Desert ecosystems: Environment and producers. Annual Review of Ecology and Systematics 4: 25-41.
- Osem Y, Perevolotsky A, and Kigel J. 2004. Grazing effect on diversity of annual plant communities in a semi-arid rangeland: Interactions with small-scale spatial and temporal variation in primary productivity. Journal of Ecology 90: 936-946.
- Pendersen BS. 1998. The role of stress in the mortality of Midwestern oaks as indicated by growth prior to death. Ecology 79: 79-93.
- Perevolotsky A. 1999. Natural conservation, reclamation, and livestock grazing in the northern Negev: Contradictory or complementary concepts? In: Hoekstra TW and Shachak M (Eds). Arid lands management: Toward ecological sustainability. Urbana, IL: University of Illinois Press.
- Puigdefábregas J. 1998. Ecological impacts of global change on drylands and their implications for desertification. Land Degradation and Development 9: 393-406.
- Ragab R and Prudhomme C. 2002. Climate change and water resources management in arid and semi-arid regions: Prospective and challenges for the 21st century. Biosystems Engineering 81: 3-34.
- Rizk AM, Hammouda FM, Ismail SE, El-Missiry MM, and Evans FJ. 1984. Irritant resiniferonol derivatives from Egyptian Thymelaea hirsuta L. Cellular and Molecular Life Sciences 40: 808-809.
- Saurez ML, Ghermandi L, and Kitzberger T. 2004. Factors predisposing episodic drought-induced tree mortality in Nothofagus – site, climatic sensitivity and growth trends. Journal of Ecology 92: 954-966.
- Schlesinger WH, Reynolds JF, Cunningham GL, Huenneke LF, Jarrell WM, Virginia RA, and Whitford WG. 1990. Biological feedbacks in global desertification. Science 247: 1043-1048.
- Seifan M, Tielbörger, K, and Kadmon R. 2010. Direct and indirect interactions among plants explain counterintuitive positive drought effects on an eastern Mediterranean shrub species. Oikos 119: 1601-1609.
- Shachak M, Boeken B, Groner E, Kadmon R, Lubin Y, Meron E, Ne’eman G, Perevolotsky A, Shkedy Y, and Ungar ED. 2008. Woody species as landscape modulators and their effect on biodiversity patterns. BioScience 58: 209-221.
- Sternberg M and Shoshani M. 2001. Influence of slope aspect on Mediterranean woody formations: Comparison of a semiarid and an arid site in Israel. Ecological Research 16: 335-345.
- Talmon Y, Sternberg M, and Grünzweig JM. 2011. Impact of rainfall manipulations and biotic controls on soil respiration in Mediterranean and desert ecosystems along an aridity gradient. Global Change Biology 17: 1108-1118.
- Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, and Bairlein F. 2002. Ecological responses to recent climate change. Nature 416: 389-395.
- Yair A and Kossovsky A. 2002. Climate and surface properties: Hydrological response of small arid and semi-arid watersheds. Geomorphology 42: 43-57.
- Yair A,and Shachak M. 1987. Studies in watershed ecology of an arid area. In: Berkofsky L and Wurtele MG (Eds). Progress in desert research. Totowa, NJ: Rowman and Littlefield.
- Zaady E, Groffman P, and Shachak M. 1996. Litter as a regulator of N and C dynamics in macrophytic patches in Negev desert soils. Soil Biology and Biochemistry 28: 39-46.




 מהדורה מודפסת
מהדורה מודפסת







